

Nonoptimal DNA topoisomerases allow maintenance of supercoiling levels and improve fitness of Streptococcus pneumoniae
- PMID: 21173181
- PMCID: PMC3067103
- DOI: 10.1128/AAC.00783-10
Nonoptimal DNA topoisomerases allow maintenance of supercoiling levels and improve fitness of Streptococcus pneumoniae
Abstract
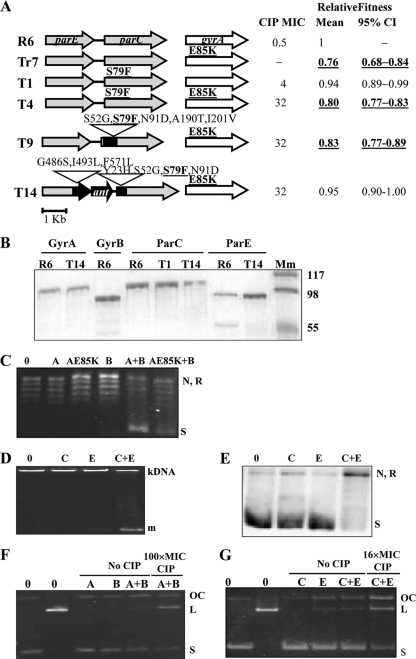
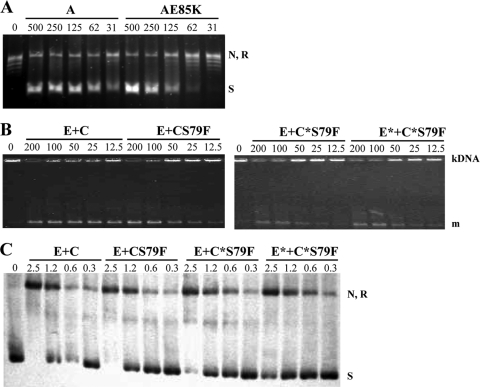
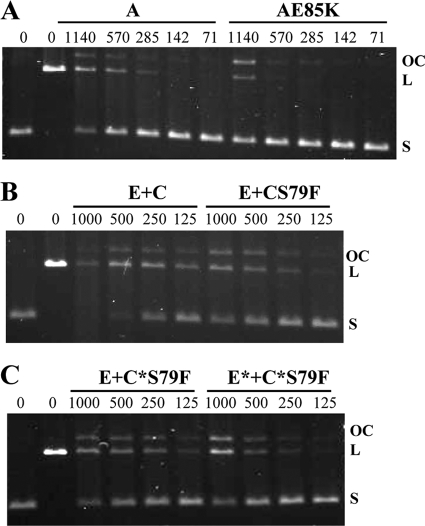
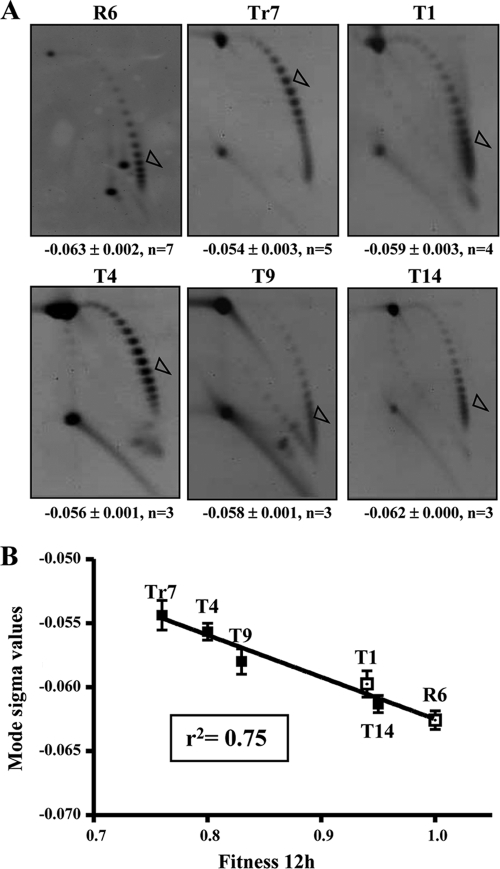
Fluoroquinolones, which target gyrase and topoisomerase IV, are used for treating Streptococcus pneumoniae infections. Fluoroquinolone resistance in this bacterium can arise via point mutation or interspecific recombination with genetically related streptococci. Our previous study on the fitness cost of resistance mutations and recombinant topoisomerases identified GyrAE85K as a high-cost change. However, this cost was compensated for by the presence of a recombinant topoisomerase IV (parC and parE recombinant genes) in strain T14. In this study, we purified wild-type and mutant topoisomerases and compared their enzymatic activities. In strain T14, both gyrase carrying GyrAE85K and recombinant topoisomerase IV showed lower activities (from 2.0- to 3.7-fold) than the wild-type enzymes. These variations of in vitro activity corresponded to changes of in vivo supercoiling levels that were analyzed by two-dimensional electrophoresis of an internal plasmid. Strains carrying GyrAE85K and nonrecombinant topoisomerases had lower (11.1% to 14.3%) supercoiling density (σ) values than the wild type. Those carrying GyrAE85K and recombinant topoisomerases showed either partial or total supercoiling level restoration, with σ values being 7.9% (recombinant ParC) and 1.6% (recombinant ParC and recombinant ParE) lower than those for the wild type. These data suggested that changes acquired by interspecific recombination might be selected because they reduce the fitness cost associated with fluoroquinolone resistance mutations. An increase in the incidence of fluoroquinolone resistance, even in the absence of further antibiotic exposure, is envisaged.
Figures
Similar articles
-
Cleavable-complex formation by wild-type and quinolone-resistant Streptococcus pneumoniae type II topoisomerases mediated by gemifloxacin and other fluoroquinolones.Antimicrob Agents Chemother. 2002 Feb;46(2):413-9. doi: 10.1128/AAC.46.2.413-419.2002. Antimicrob Agents Chemother. 2002. PMID: 11796351 Free PMC article.
-
Cloning and characterization of the parC and parE genes of Streptococcus pneumoniae encoding DNA topoisomerase IV: role in fluoroquinolone resistance.J Bacteriol. 1996 Jul;178(14):4060-9. doi: 10.1128/jb.178.14.4060-4069.1996. J Bacteriol. 1996. PMID: 8763932 Free PMC article.
-
Fitness of Streptococcus pneumoniae fluoroquinolone-resistant strains with topoisomerase IV recombinant genes.Antimicrob Agents Chemother. 2008 Mar;52(3):822-30. doi: 10.1128/AAC.00731-07. Epub 2007 Dec 26. Antimicrob Agents Chemother. 2008. PMID: 18160515 Free PMC article.
-
Mechanisms and spread of fluoroquinolone resistance in Streptococcus pneumoniae.Res Microbiol. 2000 Jul-Aug;151(6):471-3. doi: 10.1016/s0923-2508(00)00167-4. Res Microbiol. 2000. PMID: 10961461 Review.
-
Temporospatial control of topoisomerases by essential cellular processes.Curr Opin Microbiol. 2024 Dec;82:102559. doi: 10.1016/j.mib.2024.102559. Epub 2024 Nov 8. Curr Opin Microbiol. 2024. PMID: 39520813 Review.
Cited by
-
Fitness benefits in fluoroquinolone-resistant Salmonella Typhi in the absence of antimicrobial pressure.Elife. 2013 Dec 10;2:e01229. doi: 10.7554/eLife.01229. Elife. 2013. PMID: 24327559 Free PMC article.
-
Physiologic and Transcriptomic Effects Triggered by Overexpression of Wild Type and Mutant DNA Topoisomerase I in Streptococcus pneumoniae.Int J Mol Sci. 2023 Oct 31;24(21):15800. doi: 10.3390/ijms242115800. Int J Mol Sci. 2023. PMID: 37958782 Free PMC article.
-
StaR Is a Positive Regulator of Topoisomerase I Activity Involved in Supercoiling Maintenance in Streptococcus pneumoniae.Int J Mol Sci. 2023 Mar 22;24(6):5973. doi: 10.3390/ijms24065973. Int J Mol Sci. 2023. PMID: 36983048 Free PMC article.
-
Upregulation of the PatAB Transporter Confers Fluoroquinolone Resistance to Streptococcus pseudopneumoniae.Front Microbiol. 2017 Oct 26;8:2074. doi: 10.3389/fmicb.2017.02074. eCollection 2017. Front Microbiol. 2017. PMID: 29123510 Free PMC article.
-
Characterization of recombinant fluoroquinolone-resistant pneumococcus-like isolates.Antimicrob Agents Chemother. 2013 Jan;57(1):254-60. doi: 10.1128/AAC.01357-12. Epub 2012 Oct 31. Antimicrob Agents Chemother. 2013. PMID: 23114769 Free PMC article.
References
-
- Adam, H. J., D. J. Hoban, A. S. Gin, and G. G. Zhanel. 2009. Association between fluoroquinolone usage and a dramatic rise in ciprofloxacin-resistant Streptococcus pneumoniae in Canada, 1997-2006. Int. J. Antimicrob. Agents 34:82-85. - PubMed
-
- Andersson, D. I., and D. Hughes. 2010. Antibiotic resistance and its cost: is it possible to reverse resistance? Nat. Rev. Microbiol. 8:260-271. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
