

Intracellular organelles mediate cytoplasmic pulling force for centrosome centration in the Caenorhabditis elegans early embryo
- PMID: 21173218
- PMCID: PMC3017145
- DOI: 10.1073/pnas.1013275108
Intracellular organelles mediate cytoplasmic pulling force for centrosome centration in the Caenorhabditis elegans early embryo
Abstract
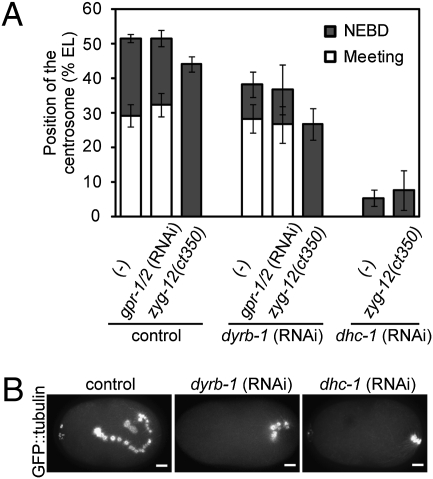
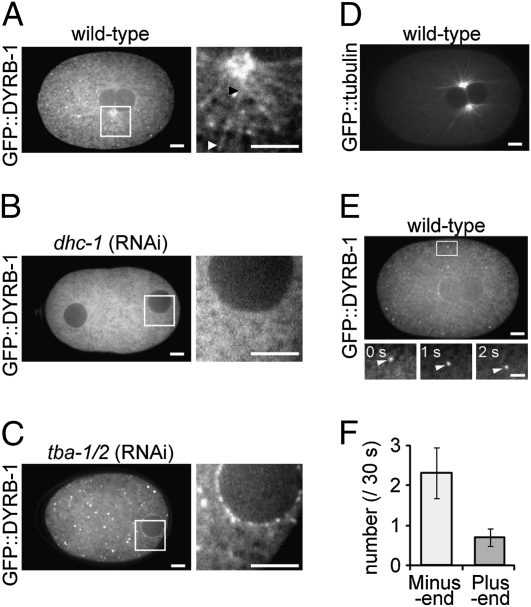
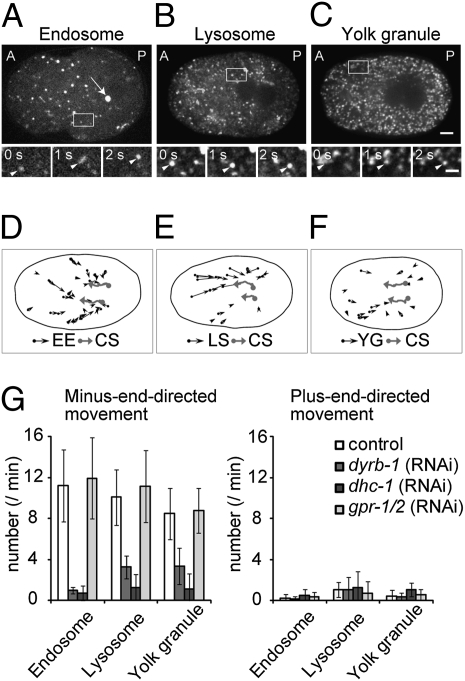
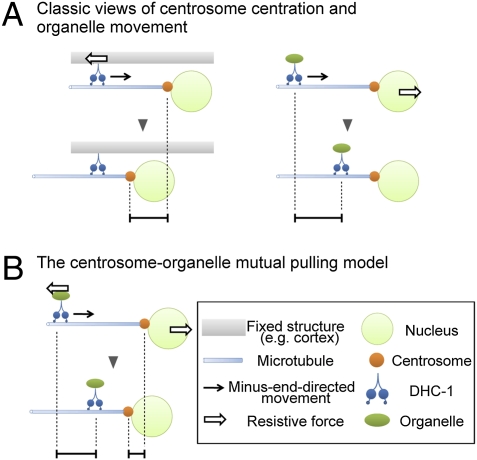
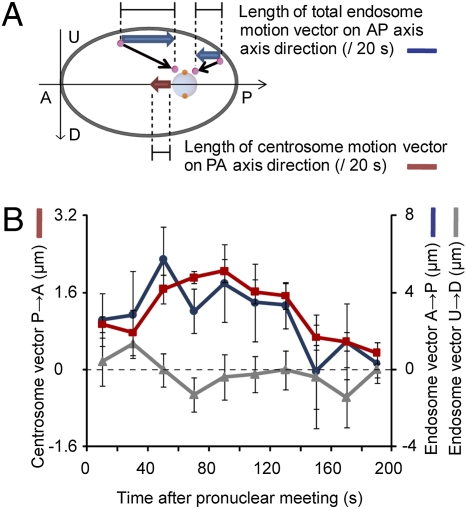
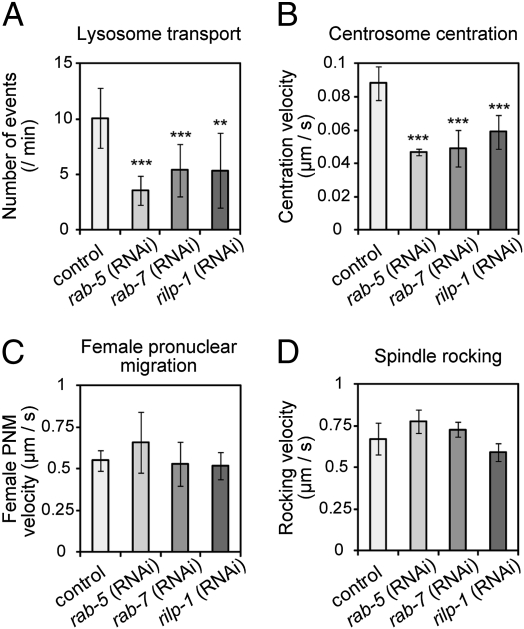
The centrosome is generally maintained at the center of the cell. In animal cells, centrosome centration is powered by the pulling force of microtubules, which is dependent on cytoplasmic dynein. However, it is unclear how dynein brings the centrosome to the cell center, i.e., which structure inside the cell functions as a substrate to anchor dynein. Here, we provide evidence that a population of dynein, which is located on intracellular organelles and is responsible for organelle transport toward the centrosome, generates the force required for centrosome centration in Caenorhabditis elegans embryos. By using the database of full-genome RNAi in C. elegans, we identified dyrb-1, a dynein light chain subunit, as a potential subunit involved in dynein anchoring for centrosome centration. DYRB-1 is required for organelle movement toward the minus end of the microtubules. The temporal correlation between centrosome centration and the net movement of organelle transport was found to be significant. Centrosome centration was impaired when Rab7 and RILP, which mediate the association between organelles and dynein in mammalian cells, were knocked down. These results indicate that minus end-directed transport of intracellular organelles along the microtubules is required for centrosome centration in C. elegans embryos. On the basis of this finding, we propose a model in which the reaction forces of organelle transport generated along microtubules act as a driving force that pulls the centrosomes toward the cell center. This is the first model, to our knowledge, providing a mechanical basis for cytoplasmic pulling force for centrosome centration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Howard J, Hyman AA. Dynamics and mechanics of the microtubule plus end. Nature. 2003;422:753–758. - PubMed
-
- Rios RM, Bornens M. The Golgi apparatus at the cell centre. Curr Opin Cell Biol. 2003;15:60–66. - PubMed
-
- Doxsey S, McCollum D, Theurkauf W. Centrosomes in cellular regulation. Annu Rev Cell Dev Biol. 2005;21:411–434. - PubMed
-
- Nigon V, Guerrier P, Monin H. L'architecture polaire de l'oeuf et les mouvements des constituants cellulaires au cours des premieres etapes du developpement chez quelques nematodes. Bull Biol Fr Belg. 1960;93:131–202.
-
- Albertson DG. Formation of the first cleavage spindle in nematode embryos. Dev Biol. 1984;101:61–72. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
