

Recruited inflammatory monocytes stimulate antiviral Th1 immunity in infected tissue
- PMID: 21173243
- PMCID: PMC3017177
- DOI: 10.1073/pnas.1005201108
Recruited inflammatory monocytes stimulate antiviral Th1 immunity in infected tissue
Abstract
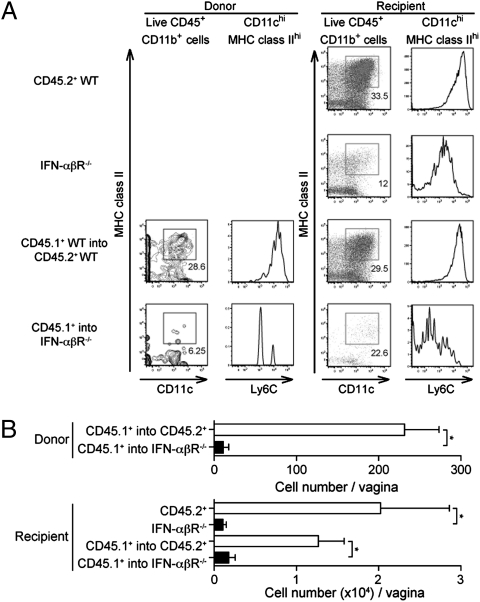
Monocytes patrol various tissues for signs of infection and inflammation. Inflammatory monocytes enter peripheral tissues at sites of microbial infection and differentiate into dendritic cells and macrophages. Here, we examined the importance of monocytes in primary mucosal infection with herpes simplex virus 2 (HSV-2), and demonstrate that monocyte-derived APCs are required to elicit IFN-γ secretion from effector Th1 cells to mediate antiviral protection. However, monocyte-derived APCs were dispensable for the generation of Th1 immunity and for the restimulation of memory Th1 cells during secondary viral challenge. These results demonstrate that distinct APC subsets are dedicated for CD4 T cell priming, elicitation, and memory recall responses to a given viral pathogen within the same mucosal tissue and reveal a specialized role for monocyte-derived APCs in the emergency response to infection.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
