

Derivation of Ca2+ signals from puff properties reveals that pathway function is robust against cell variability but sensitive for control
- PMID: 21173273
- PMCID: PMC3017167
- DOI: 10.1073/pnas.1008435108
Derivation of Ca2+ signals from puff properties reveals that pathway function is robust against cell variability but sensitive for control
Abstract
Ca(2+) is a universal second messenger in eukaryotic cells transmitting information through sequences of concentration spikes. A prominent mechanism to generate these spikes involves Ca(2+) release from the endoplasmic reticulum Ca(2+) store via inositol 1,4,5-trisphosphate (IP(3))-sensitive channels. Puffs are elemental events of IP(3)-induced Ca(2+) release through single clusters of channels. Intracellular Ca(2+) dynamics are a stochastic system, but a complete stochastic theory has not been developed yet. We formulate the theory in terms of interpuff interval and puff duration distributions because, unlike the properties of individual channels, they can be measured in vivo. Our theory reproduces the typical spectrum of Ca(2+) signals like puffs, spiking, and bursting in analytically treatable test cases as well as in more realistic simulations. We find conditions for spiking and calculate interspike interval (ISI) distributions. Signal form, average ISI and ISI distributions depend sensitively on the details of cluster properties and their spatial arrangement. In contrast to that, the relation between the average and the standard deviation of ISIs does not depend on cluster properties and cluster arrangement and is robust with respect to cell variability. It is controlled by the global feedback processes in the Ca(2+) signaling pathway (e.g., via IP(3)-3-kinase or endoplasmic reticulum depletion). That relation is essential for pathway function because it ensures frequency encoding despite the randomness of ISIs and determines the maximal spike train information content. Hence, we find a division of tasks between global feedbacks and local cluster properties that guarantees robustness of function while maintaining sensitivity of control of the average ISI.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
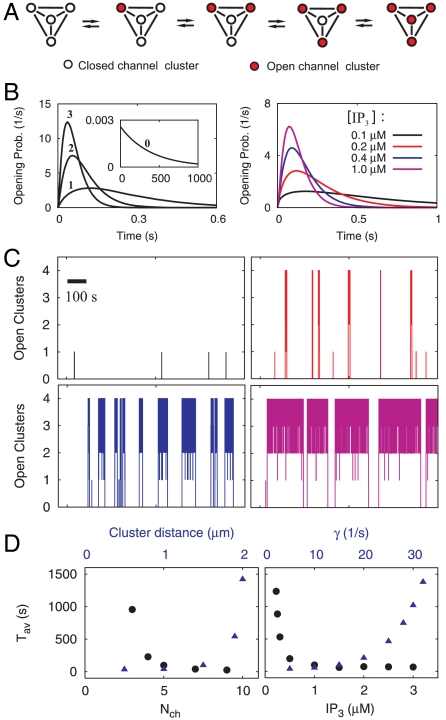
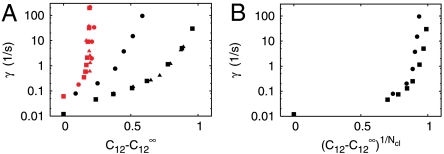
 and the channel closing rate γ between the red and black symbols. They show the long-Tav criterion for spiking Cl (red) and the short-Tav criterion Cs (black), respectively (see Conditions for Spiking and Bursting). Coupling values smaller than Cl entail essentially only local puffs, and coupling values larger than Cs cause the regime of overstimulation. The spike range increases with γ. We investigated both the four-cluster model (squares) and the eight-cluster model (circles). Cl values are similar for both models, whereas Cs values depend on the number of clusters Ncl. The spike range becomes smaller for a larger number of clusters involved in spike nucleation. Each pair of squares and circles indicates the critical (
and the channel closing rate γ between the red and black symbols. They show the long-Tav criterion for spiking Cl (red) and the short-Tav criterion Cs (black), respectively (see Conditions for Spiking and Bursting). Coupling values smaller than Cl entail essentially only local puffs, and coupling values larger than Cs cause the regime of overstimulation. The spike range increases with γ. We investigated both the four-cluster model (squares) and the eight-cluster model (circles). Cl values are similar for both models, whereas Cs values depend on the number of clusters Ncl. The spike range becomes smaller for a larger number of clusters involved in spike nucleation. Each pair of squares and circles indicates the critical ( ) at the standard parameters (Table 1), whereas coupling was changed by varying cluster distance (values range from 0.5 μm to ∞). The triangles show the critical values for the four-cluster model obtained when we varied
) at the standard parameters (Table 1), whereas coupling was changed by varying cluster distance (values range from 0.5 μm to ∞). The triangles show the critical values for the four-cluster model obtained when we varied  by changing [IP3]. That leads to very similar results. (B) The Nclth root of the short-Tav criterion Cs is shown. It suggests that Cs depends essentially exponentially on Ncl, because the Nclth root of Cs is similar for the four-cluster model (squares) and the eight-cluster model (circles).
by changing [IP3]. That leads to very similar results. (B) The Nclth root of the short-Tav criterion Cs is shown. It suggests that Cs depends essentially exponentially on Ncl, because the Nclth root of Cs is similar for the four-cluster model (squares) and the eight-cluster model (circles).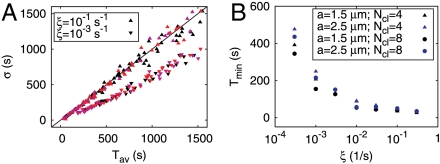
References
-
- Berridge MJ, Bootman MD, Lipp P. Calcium—A life and death signal. Nature. 1998;395:645–648. - PubMed
-
- Berridge MJ, Bootman MD, Roderick HL. Calcium signalling: Dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol. 2003;4:517–529. - PubMed
-
- Taylor CW, Swatton JE. Regulation of IP3 receptors by IP3 and Ca2+ Lect Notes Phys. 2003;623:1–16.
-
- Falcke M. Reading the patterns in living cells—The physics of Ca2+ signaling. Adv Phys. 2004;53:255–440.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
