

Hyperphagia and leptin resistance in tissue inhibitor of metalloproteinase-2 deficient mice
- PMID: 21175899
- PMCID: PMC3042479
- DOI: 10.1111/j.1365-2826.2010.02105.x
Hyperphagia and leptin resistance in tissue inhibitor of metalloproteinase-2 deficient mice
Abstract
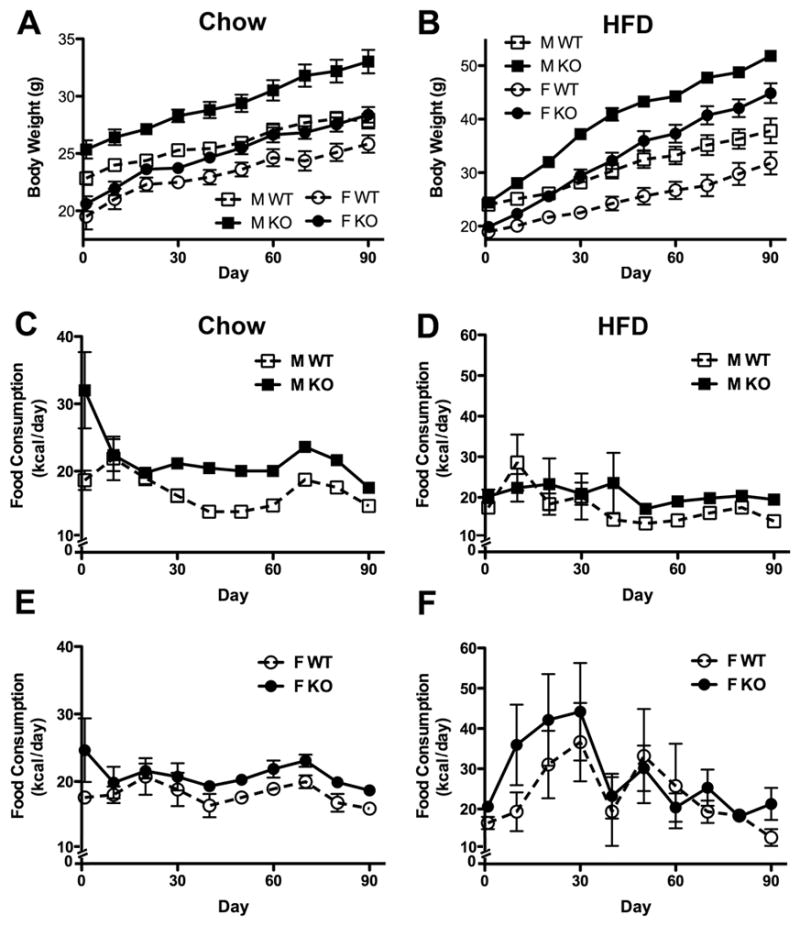
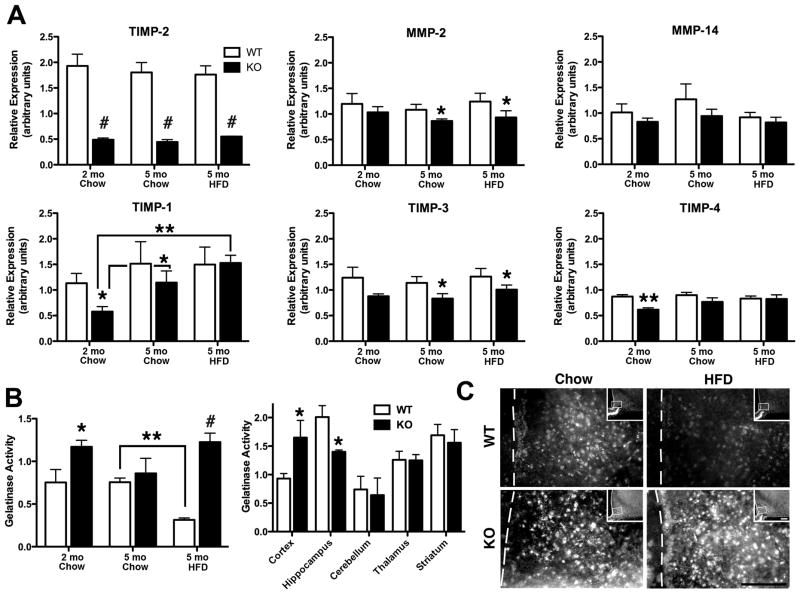
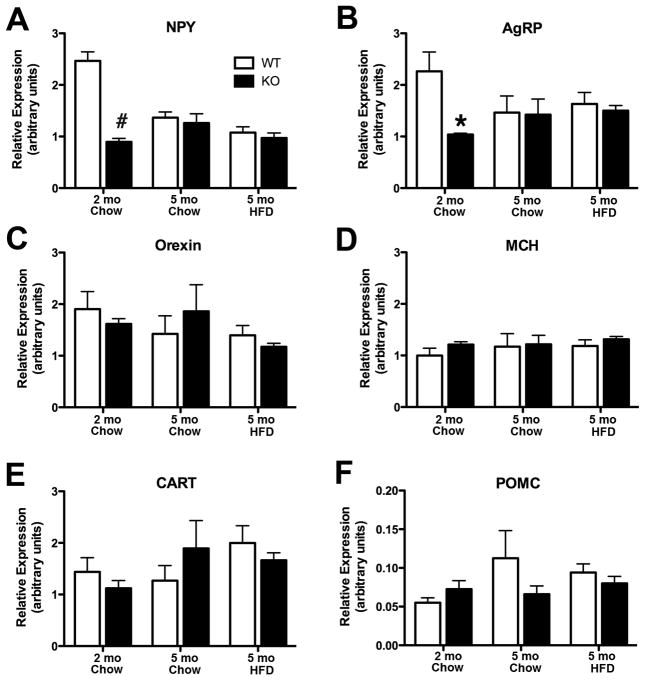
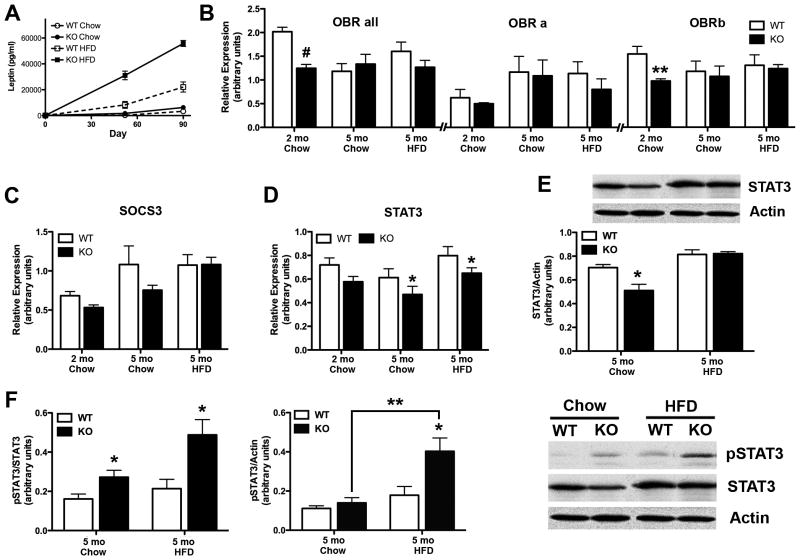
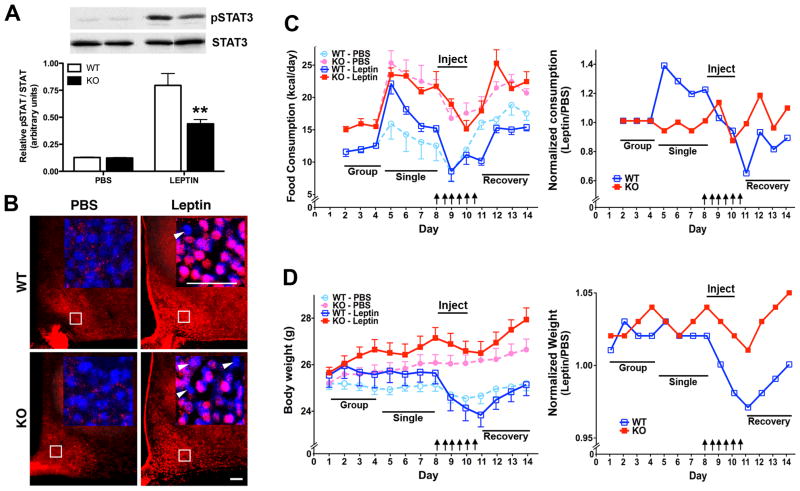
Obesity is a complex genetic and behavioural disorder arising from the improper integration of peripheral signals at central autonomic centres. For the hypothalamus to respond to dynamic physiological alterations, it must retain a degree of plasticity throughout life. Evidence is mounting that an intricate balance between matrix metalloproteinase (MMP)-mediated extracellular matrix proteolysis and tissue inhibitor of metalloproteinase (TIMP)-mediated proteolysis inhibition contributes to tissue remodelling. However, few studies have examined the role of MMPs/TIMPs in hypothalamic remodelling and energy homeostasis. To determine the contribution of TIMP-2 to the hypothalamic regulation of feeding, body mass and food consumption were monitored in TIMP-2 knockout (KO) mice fed a standard chow or high-fat diet (HFD). TIMP-2 KO mice of both sexes gained more weight than wild-type (WT) mice, even when fed the chow diet. Before the onset of obesity, TIMP-2 KO mice were hyperphagic, without increased orexigenic or decreased anorexigenic neuropeptide expression, but leptin resistant (i.e. reduced leptin-induced anorexigenic response and signal transducer and activator of transcription 3 activation). HFD exacerbated weight gain and hyperleptinaemia. In addition, proteolysis was increased in the arcuate nucleus of TIMP-2 KO mice. These data suggest a role for TIMP-2 in hypothalamic control of feeding and energy homeostasis.
© 2011 The Authors. Journal of Neuroendocrinology © 2011 Blackwell Publishing Ltd.
Conflict of interest statement
No con icts of interest, nancial or otherwise, are declared by the author(s).
Figures
References
-
- Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature. 1994;372:425–32. - PubMed
-
- Hegyi K, Fülöp K, Kovács K, Tóth S, Falus A. Leptin-induced signal transduction pathways. Cell Biol Int. 2004;28:159–69. - PubMed
-
- Valassi E, Scacchi M, Cavagnini F. Neuroendocrine control of food intake. Nutr Metab Cardiovasc Dis. 2008;18:158–68. - PubMed
-
- Farooqi IS, O’Rahilly S. Monogenic obesity in humans. Annu Rev Med. 2005;56:443–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
