

Review
doi: 10.1242/dev.046441.
Developmental regulation of axon branching in the vertebrate nervous system
Affiliations
- PMID: 21177340
- PMCID: PMC3005597
- DOI: 10.1242/dev.046441
Item in Clipboard
Review
Developmental regulation of axon branching in the vertebrate nervous system
Development.
2011 Jan.
Abstract
During nervous system development, axons generate branches to connect with multiple synaptic targets. As with axon growth and guidance, axon branching is tightly controlled in order to establish functional neural circuits, yet the mechanisms that regulate this important process are less well understood. Here, we review recent advances in the study of several common branching processes in the vertebrate nervous system. By focusing on each step in these processes we illustrate how different types of branching are regulated by extracellular cues and neural activity, and highlight some common principles that underlie the establishment of complex neural circuits in vertebrate development.
Figures
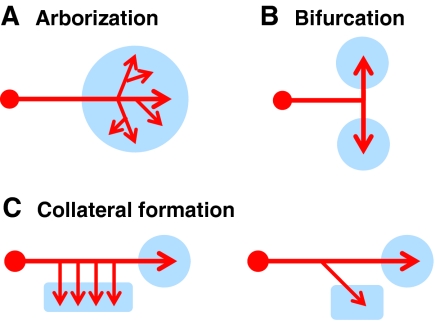
Common axon branching processes in the vertebrate nervous system. Based on morphology, complexity and function, axon branching is grouped into three classes in this review: arborization, bifurcation and collateral formation. (A) Arborization usually occurs at axon terminals in the target region (blue circle), where most elaborated branched structures are generated by repetitive branch formation. (B) Axon bifurcation also occurs at axon terminals, but tends to generate two daughter branches that project to targets (blue circles) in opposite directions. (C) In collateral formation, daughter branches sprout from the middle of axonal shafts, away from axon terminals, and innervate targets that are usually different from the main axons (blue rectangles versus circles). In some axons, only one collateral branch forms (right), but in others multiple collaterals can form to synapse with similar targets (left). Structurally, collateral formation might be more closely related to axon bifurcation, as only the first-order branches are generated. The main features that distinguish them are the functions of the daughter branches and the angles between the daughter branch and the main axon.
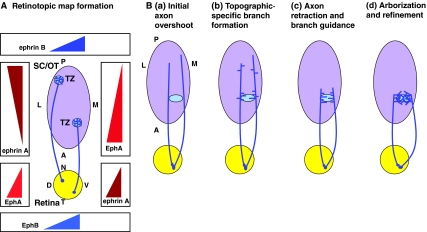
Development of topographic-specific axonal branches in the visual system. (A) Retinotopic map formation is regulated by the interactions of ephrin A/EphA gradients. To preserve the spatial information received in the retina (yellow circle), the retinal ganglion cell (RGC) axons project to the terminal zone (TZ, blue circle) of their cortical target (purple oval) in a topographic fashion to form the retinotopic map. In mammals, the target is the superior colliculus (SC), whereas in Xenopus, chick and zebrafish it is the optic tectum (OT). The retinotopic map forms via the repulsive interaction of ephrin A, which is expressed in the target structure, and the EphA receptors, which are expressed on growing RGC axons. As shown in the left gradients, EphAs are expressed in an increasing nasal-temporal (N-T) gradient in the retina, whereas ephrin A ligands are expressed in an increasing anterior-posterior (A-P) gradient in the cortical target. Other signaling gradients, including ephrin A/EphA reverse signaling (right-hand side), as well as ephrin B/EphB (top and bottom), are also shown here and contribute to RGC axon branching. (B) RGC terminal arbor development occurs via multiple steps in chick and mouse embryos. (a) RGC axons initially overshoot the correct TZ and interstitial branches form along the A-P axis of the axon. (b) Branch formation increases at the correct topographic-specific TZ, a process that is regulated by opposing ephrin A/EphA gradients and by global expression of brain-derived neurotrophic factor (BDNF). (c) The overextended axons then retract to the correct TZ, and branches are preferentially guided to the TZ, possibly via an EphB gradient on the retinal dorsal-ventral (D-V) axis and an ephrin B gradient on the lateral-medial (L-M) axis of the target. (d) An activity-dependent process then eliminates branches outside of the correct TZ while retaining correctly positioned branches. Figures are modified with permission (McLaughlin and O'Leary, 2005).
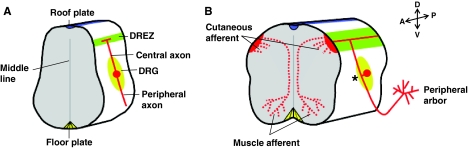
Development of sensory axon branching in the spinal cord. Schematized cross-sections of a vertebrate developing spinal cord illustrate three branching forms of dorsal root ganglion (DRG) sensory axons (Davis et al., 1989; Mendelson et al., 1992; Mirnics and Koerber, 1995a; Mirnics and Koerber, 1995b; Ozaki and Snider, 1997; Ramon y Cajal, 1904). (A) The DRG flanks the spinal cord and contains the cell bodies (red circle) of sensory neurons that initially generate two axons. One axon (the peripheral axon) projects to the skin or muscle, whereas the other axon (the central axon) projects centrally to the spinal cord. (B) Later in development, these two axons fuse (asterisk) to form a single axon and establish a pseudo-unipolar morphology (Ramon y Cajal, 1904). The peripheral axons arborize in their targets to form the peripheral arbors, while the central axons bifurcate once they reach the dorsal root entry zone (DREZ; green stripe) of the spinal cord and continue to extend in opposite directions along the anterior-posterior (A-P) axis. There are two functional significances of this bifurcation: it allows sensory information to be transmitted to the high-order relay neurons in the spinal cord or the brain stem via its ascending projections (growing anteriorly); and, both the ascending and descending (growing anteriorly and posteriorly) axons sprout interstitial collateral branches that invade the gray matter of the spinal cord. The collaterals of cutaneous and motor afferents terminate at different lamina on the dorsal-ventral (D-V) axis of the spinal cord and form monosynaptic or polysynaptic reflex circuits. The terminals of these collaterals also arborize, as shown here for the muscle afferents.
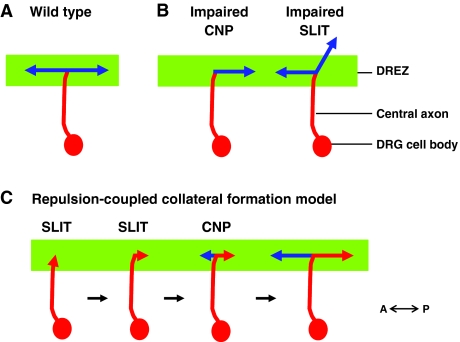
An illustration of sensory axon bifurcation. A schematic of sensory axon bifurcation in the dorsal root entry zone (DREZ) of the mouse spinal cord. (A) In wild-type mouse embryos, the central axon (red) of the dorsal root ganglion (DRG) bifurcates at the DREZ (green stripe) of the spinal cord. The two resulting branches (blue) extend in opposite directions along the anterior-posterior (A-P) axis, perpendicular to the primary axon (red). (B) Summary of bifurcation defects in mouse mutants. Mice that lack C-type natriuretic peptide (CNP; also known as NPPC) signaling include: a spontaneous Nppc mutant [lbab (long bone abnormality)]; a targeted Nppc knockout; a spontaneous natriuretic peptide receptor 2 (Npr2) mutant; and a targeted deletion of cGMP-dependent protein kinase GI (Prkg1) (Schmidt et al., 2009; Schmidt et al., 2007; Zhao and Ma, 2009; Zhao et al., 2009). In these mutants, DRG axons fail to bifurcate in the DREZ, resulting in the loss of the second branch. In mutants with impaired SLIT signaling, such as in Slit1;Slit2 or Robo1;Robo2 double mutants, DRG axons still bifurcate, but one daughter branch is misguided and enters the spinal cord (Ma and Tessier-Lavigne, 2007). (C) A repulsion-coupled collateral formation model that describes the sequential steps required for sensory axon bifurcation in the spinal cord. After the primary sensory axon (red) first reaches the DREZ (green stripe), it encounters SLIT proteins, which are present next to the DREZ. These cues stop the axon and guide it to make a turn randomly to the A-P track of the DREZ. Immediately after turning, the axon receives an instructive signal from CNP and makes a new branch (blue), which sprouts as a collateral from the turning site and is biochemically different from the other branch (red).
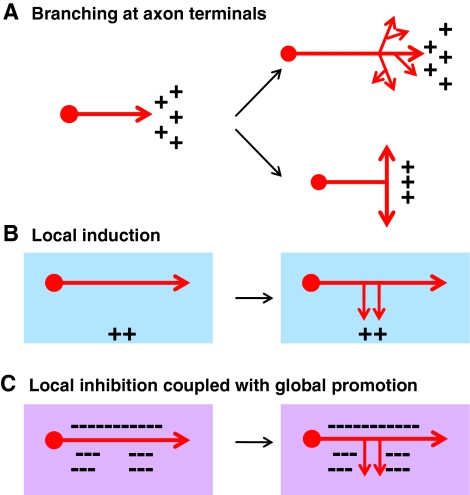
Mechanisms that specify the location of axon branching. (A) Branches often occur at the end of axonal terminals. The presence of branch-promoting factors (represented by ++), such as nerve growth factor (NGF), wingless-related MMTV integration site (Wnt) factors and C-type natriuretic peptide (CNP), in the target region, can promote the formation of both terminal arbors (top) and bifurcation (bottom). However, the mechanisms that underlie these different types of branching remain to be determined. (B) Collateral branches can be promoted by a positive cue interstitially along the axon. When navigating to find their targets during early development, many axons either cannot form branches or are inhibited by the local environment (blue box). A positive branch-promoting cue (++) present in the target area can stimulate the formation of an interstitial branch at a specific location along the axon and promote its growth to become a collateral branch. (C) A mechanism involving local inhibition (represented by dashed line) coupled with global activation can also specify the location of interstitial branch formation along the axon. In this mechanism, branch formation can occur anywhere along the axon, either by an intrinsic mechanism or by a positive branch-promoting factor that is present along the route of axonal growth (purple box). Inhibitory factors, such as ephrin A/EphA, present near the axons specify branching location by restricting branch formation to certain regions of the axon.
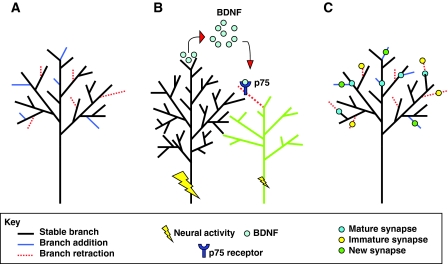
Mechanisms underlying activity-dependent regulation of axon branching. (A) Axon branching often occurs as a dynamic process that involves branch addition and branch retraction. Branching dynamics can be modulated by neural activity to produce a bias toward branch addition or retraction, leading to a net change in the arbor morphology. (B) Neighboring axonal branches compete for innervation territory in the target structure. Typically, the axon experiencing the greatest amount of activity `wins' the competition by inhibiting the arborization of neighboring axons. In sympathetic neurons, this phenomenon might be mediated by a brain-derived neurotrophic factor (BDNF)-nerve growth factor receptor (p75) interaction, in which the `winning' branch secretes BDNF that then binds to the p75 receptor on the `losing' axon, resulting in the loss of that branch due to axon degeneration (illustrated here as branch retraction for simplicity). (C) Axon branching is tightly coupled to synapse development. The maturity of new synapses is a significant criterion for determining which branches will be retained or retracted, in addition to where new branches will form. Mature synapses can halt branch retraction, whereas immature synapses are associated with retracting branches. New synapses preferentially form on new branches, and new branches preferentially form near mature synapses.
References
-
- Acebes A., Ferrus A. (2000). Cellular and molecular features of axon collaterals and dendrites. Trends Neurosci. 23, 557-565 - PubMed
-
- Alexander M., Selman G., Seetharaman A., Chan K. K., D'Souza S. A., Byrne A. B., Roy P. J. (2010). MADD-2, a homolog of the Opitz syndrome protein MID1, regulates guidance to the midline through UNC-40 in Caenorhabditis elegans. Dev. Cell 18, 961-972 - PubMed
-
- Alsina B., Vu T., Cohen-Cory S. (2001). Visualizing synapse formation in arborizing optic axons in vivo: dynamics and modulation by BDNF. Nat. Neurosci. 4, 1093-1101 - PubMed
-
- Anitha A., Nakamura K., Yamada K., Suda S., Thanseem I., Tsujii M., Iwayama Y., Hattori E., Toyota T., Miyachi T., et al. (2008). Genetic analyses of roundabout (ROBO) axon guidance receptors in autism. Am. J. Med. Genet. B Neuropsychiatr. Genet. 147, 1019-1027 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
