

Genome-wide evidence for an essential role of the human Staf/ZNF143 transcription factor in bidirectional transcription
- PMID: 21177654
- PMCID: PMC3082894
- DOI: 10.1093/nar/gkq1301
Genome-wide evidence for an essential role of the human Staf/ZNF143 transcription factor in bidirectional transcription
Abstract
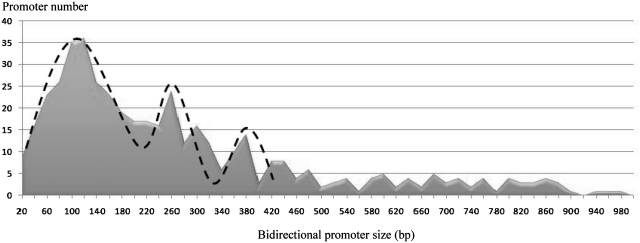
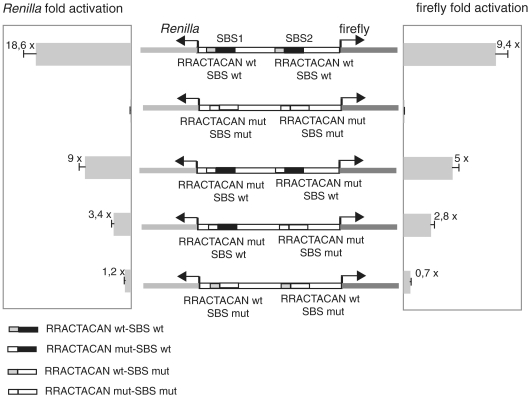
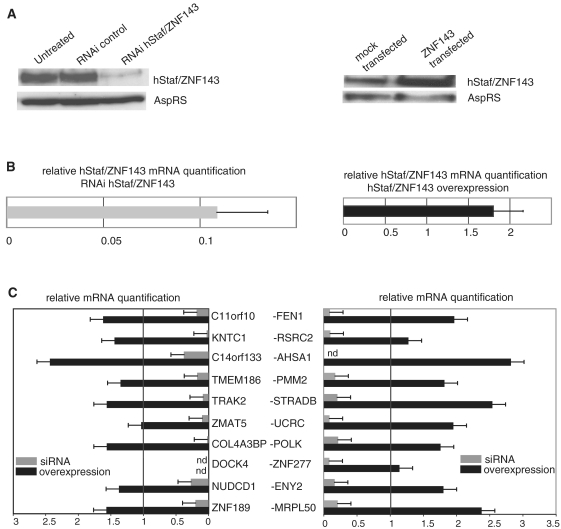
In the human genome, ∼ 10% of the genes are arranged head to head so that their transcription start sites reside within <1 kbp on opposite strands. In this configuration, a bidirectional promoter generally drives expression of the two genes. How bidirectional expression is performed from these particular promoters constitutes a puzzling question. Here, by a combination of in silico and biochemical approaches, we demonstrate that hStaf/ZNF143 is involved in controlling expression from a subset of divergent gene pairs. The binding sites for hStaf/ZNF143 (SBS) are overrepresented in bidirectional versus unidirectional promoters. Chromatin immunoprecipitation assays with a significant set of bidirectional promoters containing putative SBS revealed that 93% of them are associated with hStaf/ZNF143. Expression of dual reporter genes directed by bidirectional promoters are dependent on the SBS integrity and requires hStaf/ZNF143. Furthermore, in some cases, functional SBS are located in bidirectional promoters of gene pairs encoding a noncoding RNA and a protein gene. Remarkably, hStaf/ZNF143 per se exhibits an inherently bidirectional transcription activity, and together our data provide the demonstration that hStaf/ZNF143 is indeed a transcription factor controlling the expression of divergent protein-protein and protein-non-coding RNA gene pairs.
Figures


References
-
- Adachi N, Lieber MR. Bidirectional gene organization: a common architectural feature of the human genome. Cell. 2002;109:807–809. - PubMed
-
- Koyanagi KO, Hagiwara M, Itoh T, Gojobori T, Imanishi T. Comparative genomics of bidirectional gene pairs and its implications for the evolution of a transcriptional regulation system. Gene. 2005;353:169–176. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
