

The biological control of voluntary exercise, spontaneous physical activity and daily energy expenditure in relation to obesity: human and rodent perspectives
- PMID: 21177942
- PMCID: PMC3008631
- DOI: 10.1242/jeb.048397
The biological control of voluntary exercise, spontaneous physical activity and daily energy expenditure in relation to obesity: human and rodent perspectives
Abstract
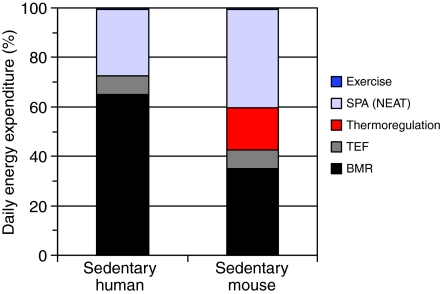
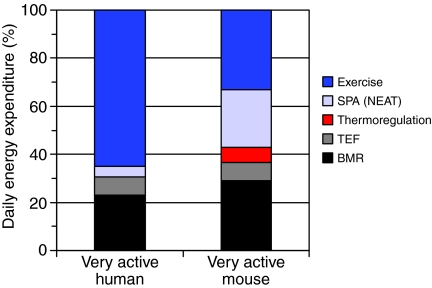
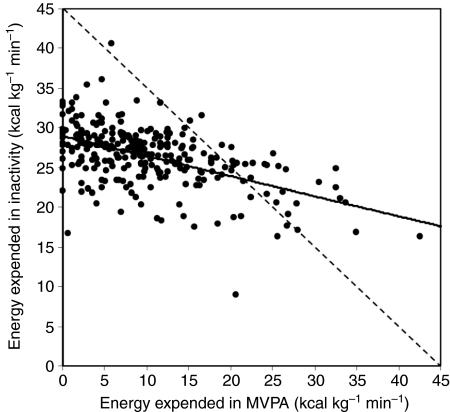
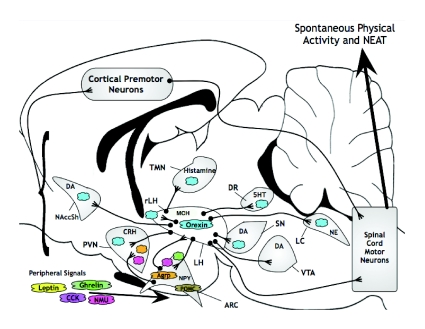
Mammals expend energy in many ways, including basic cellular maintenance and repair, digestion, thermoregulation, locomotion, growth and reproduction. These processes can vary tremendously among species and individuals, potentially leading to large variation in daily energy expenditure (DEE). Locomotor energy costs can be substantial for large-bodied species and those with high-activity lifestyles. For humans in industrialized societies, locomotion necessary for daily activities is often relatively low, so it has been presumed that activity energy expenditure and DEE are lower than in our ancestors. Whether this is true and has contributed to a rise in obesity is controversial. In humans, much attention has centered on spontaneous physical activity (SPA) or non-exercise activity thermogenesis (NEAT), the latter sometimes defined so broadly as to include all energy expended due to activity, exclusive of volitional exercise. Given that most people in Western societies engage in little voluntary exercise, increasing NEAT may be an effective way to maintain DEE and combat overweight and obesity. One way to promote NEAT is to decrease the amount of time spent on sedentary behaviours (e.g. watching television). The effects of voluntary exercise on other components of physical activity are highly variable in humans, partly as a function of age, and have rarely been studied in rodents. However, most rodent studies indicate that food consumption increases in the presence of wheels; therefore, other aspects of physical activity are not reduced enough to compensate for the energetic cost of wheel running. Most rodent studies also show negative effects of wheel access on body fat, especially in males. Sedentary behaviours per se have not been studied in rodents in relation to obesity. Several lines of evidence demonstrate the important role of dopamine, in addition to other neural signaling networks (e.g. the endocannabinoid system), in the control of voluntary exercise. A largely separate literature points to a key role for orexins in SPA and NEAT. Brain reward centers are involved in both types of physical activities and eating behaviours, likely leading to complex interactions. Moreover, voluntary exercise and, possibly, eating can be addictive. A growing body of research considers the relationships between personality traits and physical activity, appetite, obesity and other aspects of physical and mental health. Future studies should explore the neurobiology, endocrinology and genetics of physical activity and sedentary behaviour by examining key brain areas, neurotransmitters and hormones involved in motivation, reward and/or the regulation of energy balance.
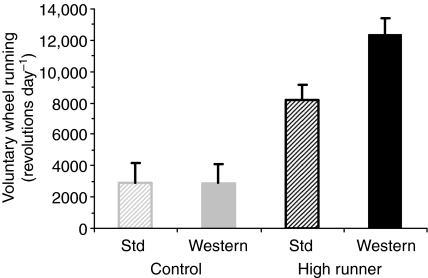
Figures

References
-
- Ahlenius S., Hillegaart V., Thorell G., Magnusson O., Fowler C. J. (1987). Suppression of exploratory locomotor activity and increase in dopamine turnover following the local application of cis-flupenthixol into limbic projection areas of the rat striatum. Brain Res. 402, 131-138 - PubMed
-
- Aidman E. V., Woollard S. (2003). The influence of self-reported exercise addiction on acute emotional and physiological responses to brief exercise deprivation. Psychol. Sport Exerc. 4, 225-226
-
- Ainsworth B. E., Haskell W. L., Whitt M. C., Irwin M. L., Swartz A. M., Strath S. J., O’Brien W. L., Bassett D. R., Jr, Schmitz K. H., Emplaincourt P. O., et al. (2000). Compendium of physical activities: an update of activity codes and MET intensities. Med. Sci. Sports Exerc. 32, S498-S504 - PubMed
-
- Allegre B., Souville M., Therma P., Griffiths M. (2006). Definitions and measures of exercise dependance. Addict. Res. Theory. 14, 631-646
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
