

Genome-wide analysis of alternative splicing in Caenorhabditis elegans
- PMID: 21177968
- PMCID: PMC3032936
- DOI: 10.1101/gr.114645.110
Genome-wide analysis of alternative splicing in Caenorhabditis elegans
Abstract
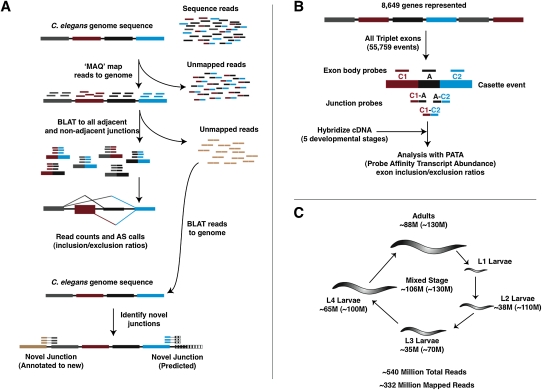
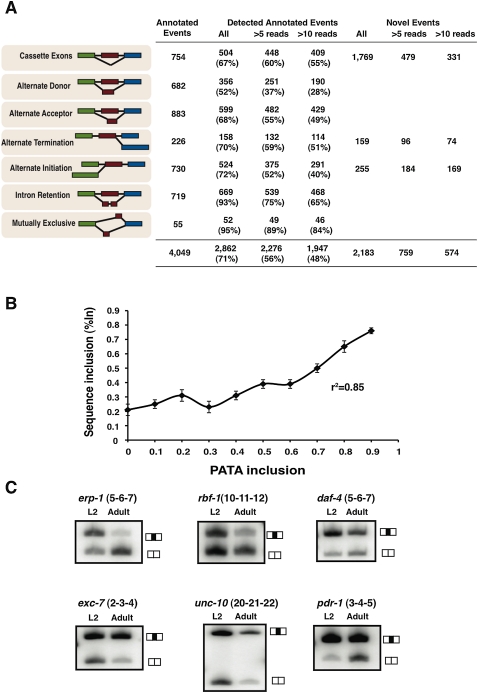
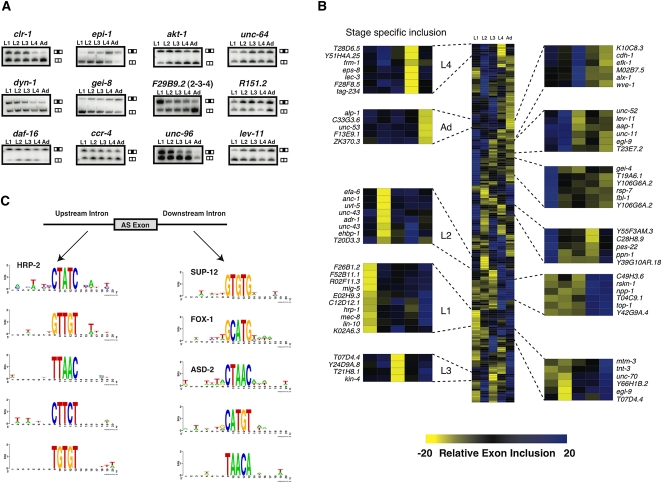
Alternative splicing (AS) plays a crucial role in the diversification of gene function and regulation. Consequently, the systematic identification and characterization of temporally regulated splice variants is of critical importance to understanding animal development. We have used high-throughput RNA sequencing and microarray profiling to analyze AS in C. elegans across various stages of development. This analysis identified thousands of novel splicing events, including hundreds of developmentally regulated AS events. To make these data easily accessible and informative, we constructed the C. elegans Splice Browser, a web resource in which researchers can mine AS events of interest and retrieve information about their relative levels and regulation across development. The data presented in this study, along with the Splice Browser, provide the most comprehensive set of annotated splice variants in C. elegans to date, and are therefore expected to facilitate focused, high resolution in vivo functional assays of AS function.
Figures
References
-
- Barash Y, Bejerano G, Friedman N 2001. A simple hyper-geometric approach for discovering putative transcription factor binding sites. Lect Notes Comput Sci 2149: 278–293
-
- Blencowe BJ 2006. Alternative splicing: New insights from global analyses. Cell 126: 37–47 - PubMed
-
- Calarco JA, Superina S, O'Hanlon D, Gabut M, Raj B, Pan Q, Skalska U, Clarke L, Gelinas D, van der Kooy D, et al. 2009. Regulation of vertebrate nervous system alternative splicing and development by an SR-related protein. Cell 138: 898–910 - PubMed
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials