

Alterations of action potentials and the localization of Nav1.6 sodium channels in spared axons after hemisection injury of the spinal cord in adult rats
- PMID: 21177993
- PMCID: PMC3074424
- DOI: 10.1152/jn.00810.2010
Alterations of action potentials and the localization of Nav1.6 sodium channels in spared axons after hemisection injury of the spinal cord in adult rats
Abstract
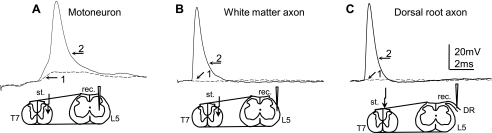
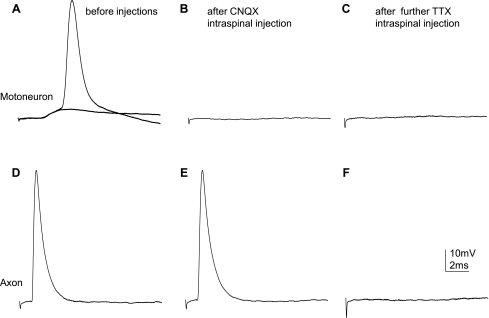
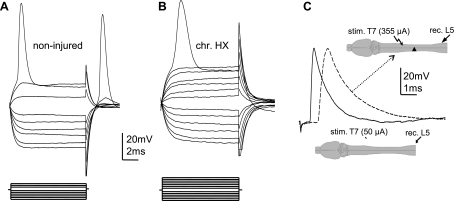
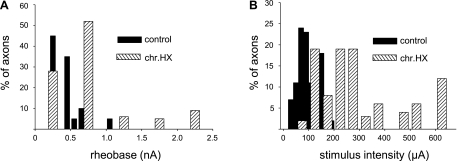
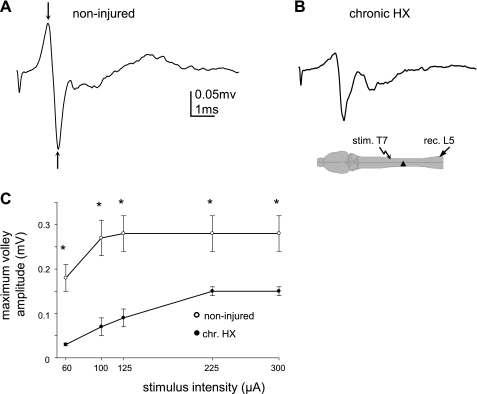
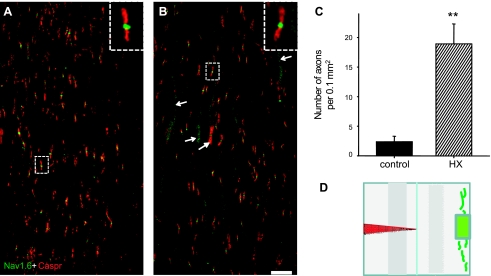
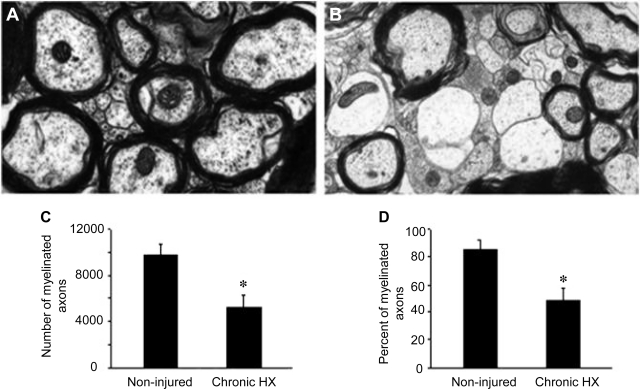
Previously, we reported a pronounced reduction in transmission through surviving axons contralateral to chronic hemisection (HX) of adult rat spinal cord. To examine the cellular and molecular mechanisms responsible for this diminished transmission, we recorded intracellularly from lumbar lateral white matter axons in deeply anesthetized adult rats in vivo and measured the propagation of action potentials (APs) through rubrospinal/reticulospinal tract (RST/RtST) axons contralateral to chronic HX at T10. We found decreased excitability in these axons, manifested by an increased rheobase to trigger APs and longer latency for AP propagation passing the injury level, without significant differences in axonal resting membrane potential and input resistance. These electrophysiological changes were associated with altered spatial localization of Nav1.6 sodium channels along axons: a subset of axons contralateral to the injury exhibited a diffuse localization (>10 μm spread) of Nav1.6 channels, a pattern characteristic of demyelinated axons (Craner MJ, Newcombe J, Black JA, Hartle C, Cuzner ML, Waxman SG. Proc Natl Acad Sci USA 101: 8168-8173, 2004b). This result was substantiated by ultrastructural changes seen with electron microscopy, in which an increased number of large-caliber, demyelinated RST axons were found contralateral to the chronic HX. Therefore, an increased rheobase, pathological changes in the distribution of Nav1.6 sodium channels, and the demyelination of contralateral RST axons are likely responsible for their decreased conduction chronically after HX and thus may provide novel targets for strategies to improve function following incomplete spinal cord injury.
Figures
References
-
- Arvanian VL, Mendell LM. Removal of NMDA receptor Mg2+ block extends the action of NT-3 on synaptic transmission in neonatal rat motoneurons. J Neurophysiol 86: 123–129, 2001 - PubMed
-
- Ballermann M, Fouad K. Spontaneous locomotor recovery in spinal cord injured rats is accompanied by anatomical plasticity of reticulospinal fibers. Eur J Neurosci 23: 1988–1996, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous