

Single-gene tuning of Caulobacter cell cycle period and noise, swarming motility, and surface adhesion
- PMID: 21179017
- PMCID: PMC3018171
- DOI: 10.1038/msb.2010.95
Single-gene tuning of Caulobacter cell cycle period and noise, swarming motility, and surface adhesion
Abstract
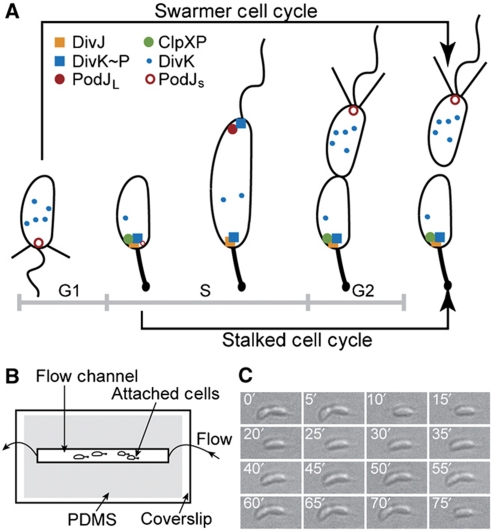
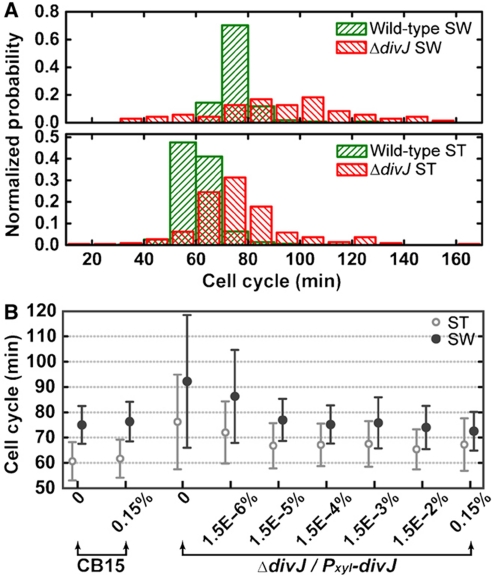
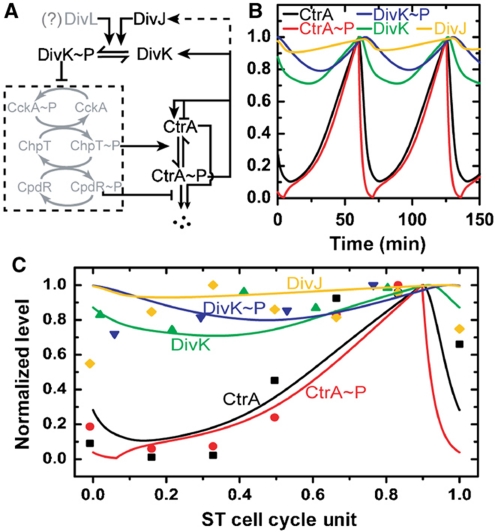
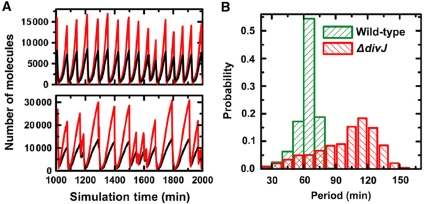
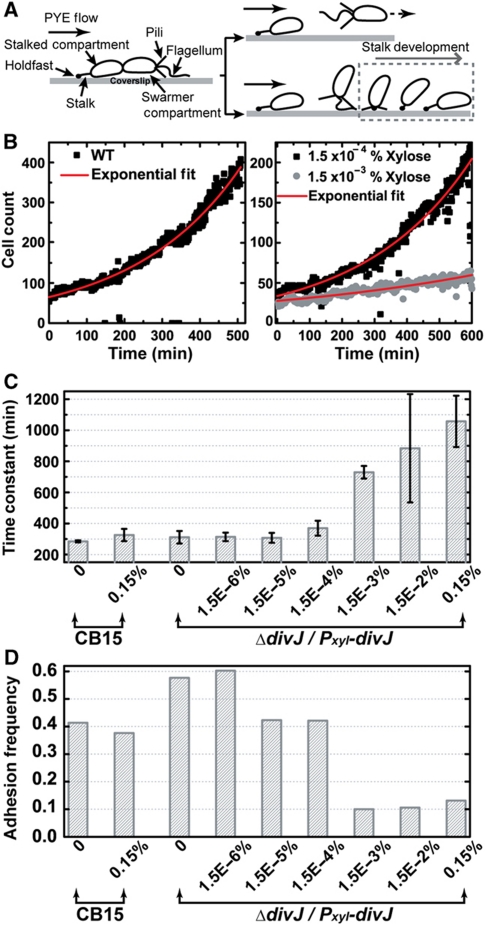
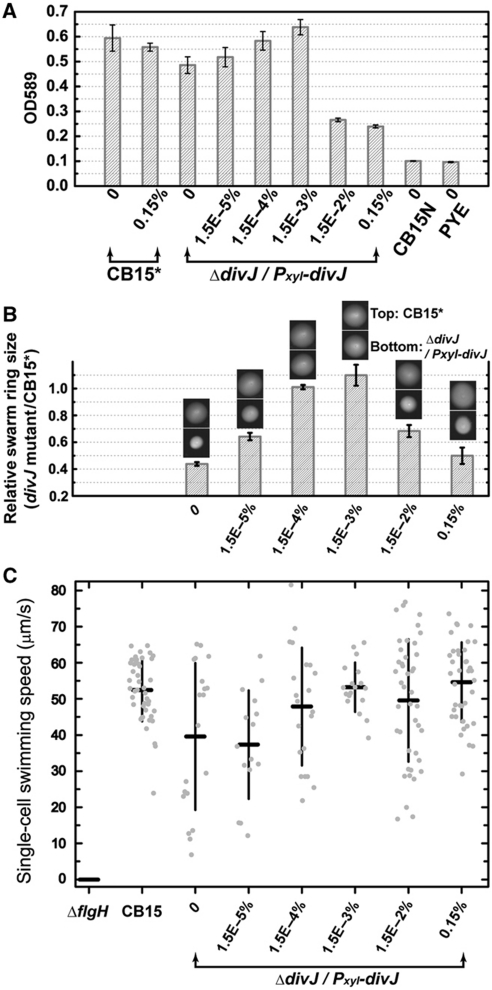
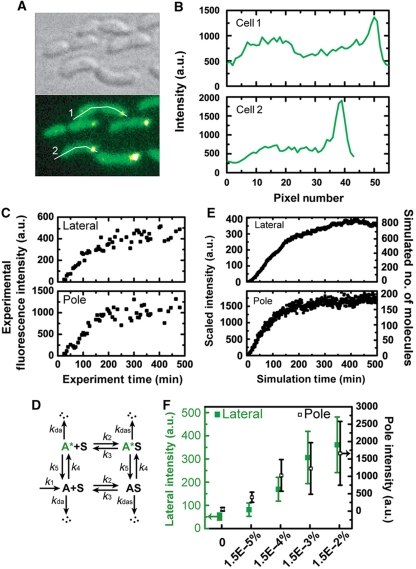
Sensor histidine kinases underlie the regulation of a range of physiological processes in bacterial cells, from chemotaxis to cell division. In the gram-negative bacterium Caulobacter crescentus, the membrane-bound histidine kinase, DivJ, is a polar-localized regulator of cell cycle progression and development. We show that DivJ localizes to the cell pole through a dynamic diffusion and capture mechanism rather than by active localization. Analysis of single C. crescentus cells in microfluidic culture demonstrates that controlled expression of divJ permits facile tuning of both the mean and noise of the cell division period. Simulations of the cell cycle that use a simplified protein interaction network capture previously measured oscillatory protein profiles, and recapitulate the experimental observation that deletion of divJ increases the cell cycle period and noise. We further demonstrate that surface adhesion and swarming motility of C. crescentus in semi-solid media can also be tuned by divJ expression. We propose a model in which pleiotropic control of polar cell development by the DivJ-DivK-PleC signaling pathway underlies divJ-dependent tuning of cell swarming and adhesion behaviors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Alley MRK, Maddock JR, Shapiro L (1992) Polar localization of a bacterial chemreceptor. Genes Dev 6: 825–836 - PubMed
-
- Bardy SL, Maddock JR (2007) Polar explorations: recent insights into the polarity of bacterial proteins. Curr Opin Microbiol 10: 617–623 - PubMed
-
- Battogtokh D, Aihara K, Tyson JJ (2006) Synchronization of eukaryotic cells by periodic forcing. Phys Rev Lett 96: 148102–148104 - PubMed
-
- Biondi EG, Reisinger SJ, Skerker JM, Arif M, Perchuk BS, Ryan KR, Laub MT (2006) Regulation of the bacterial cell cycle by an integrated genetic circuit. Nature 444: 899–904 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
