

Review
doi: 10.1038/nrm3029.
Inventory control: cytochrome c oxidase assembly regulates mitochondrial translation
Affiliations
- PMID: 21179059
- PMCID: PMC3376542
- DOI: 10.1038/nrm3029
Item in Clipboard
Review
Inventory control: cytochrome c oxidase assembly regulates mitochondrial translation
Nat Rev Mol Cell Biol.
2011 Jan.
Abstract
Mitochondria maintain genome and translation machinery to synthesize a small subset of subunits of the oxidative phosphorylation system. To build up functional enzymes, these organellar gene products must assemble with imported subunits that are encoded in the nucleus. New findings on the early steps of cytochrome c oxidase assembly reveal how the mitochondrial translation of its core component, cytochrome c oxidase subunit 1 (Cox1), is directly coupled to the assembly of this respiratory complex.
Figures
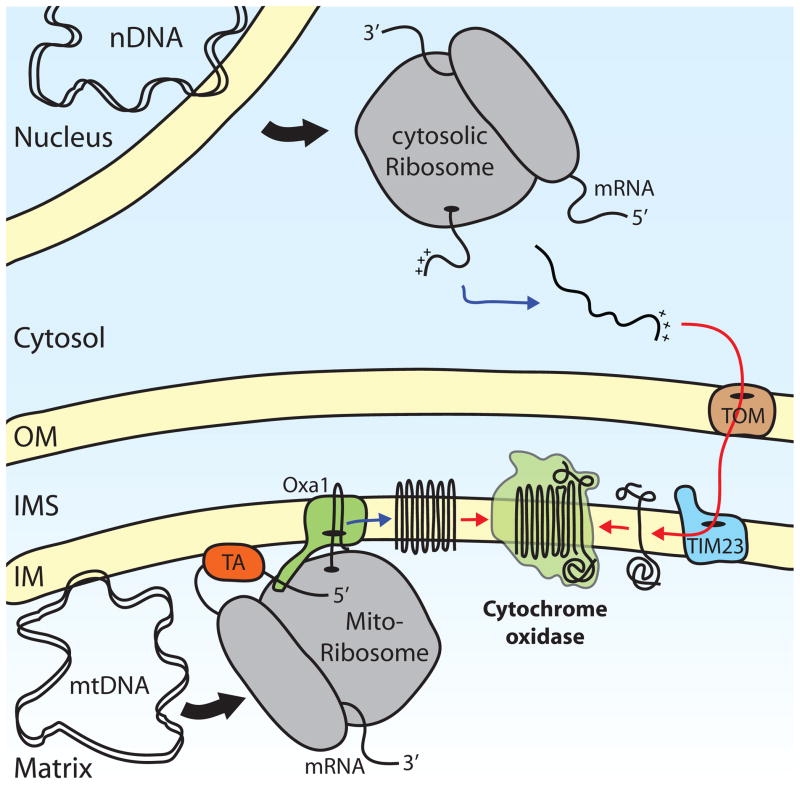
Respiratory chain subunits are encoded by both nuclear DNA (nDNA), such as Cox5 and mitochondrial DNA (mtDNA), such as Cox1. Nuclear-encoded proteins are translated on cytosolic ribosomes as precursor proteins that contain information that targets them for mitochondria (+++). Precursors are recognized and transported across the mitochondrial membranes by the translocase of the outer membrane (TOM) complex and the translocase of the inner membrane (TIM23) complex, which cooperate in this process. mtDNA is associated with the inner mitochondrial membrane (IM) and its transcription, as well as translation, occurs in close proximity. Mitochondrial ribosomes translate mRNAs that are bound to the membrane by specific translational activators (TA), such as Pet309 in the case of the mitochondria-encoded Cox1. The translocase Oxa1 inserts nascent polypeptides in a co-translational manner into the membrane and helps these proteins to attain their correct topology. After the insertion of both mitochondria- and nuclear-encoded proteins into the membrane, these subunits can assemble into a functional complex. (OM, outer mitochondrial membrane; IMS, intermembrane space)
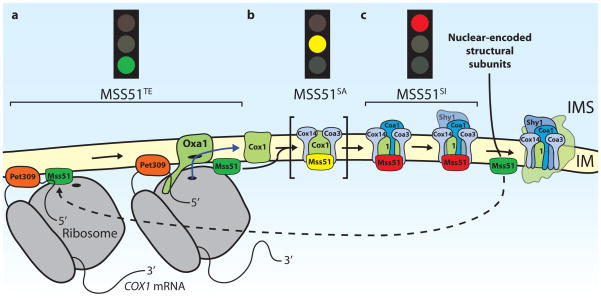
Mss51 regulates the translation of Cox1 by interacting with Cox1 mRNA, and also Cox1 protein that has not yet been assembled into the mature cytochrome oxidase complex. a| Cox1 is synthesized by mitochondrial ribosomes upon activation by the translational activators Pet309 and Mss51. Here Mss51 is in a t ranslation-e ffective state (MSS51TE). Cox1 is inserted into the inner mitochondrial membrane (IM) co-translationally by the Oxa1 translocase. b| Mss51 is also required for Cox1 translation independently of the 5′UTR. It interacts with newly synthesized Cox1 and the transmembrane proteins Cox14 and Coa3 (c ytochrome o xidase a ssembly), which promote the Cox1–Mss51 interaction. In this s equestered a ctive state (MSS51SA), which has been observed in cells lacking the assembly factor Coa1, Mss51 may be able to initiate further rounds of Cox1 synthesis on mitochondrial ribosomes (not shown) or it might be loosely associated with this complex so that some Mss51 can be released to activate translation. c| The association of Coa1 with this complex in wild type cells converts Mss51 to a s equestered i nactive state (MSS51SI), which prevents it from activating translation. Moreover, Coa1 association promotes the binding of Shy1 to Cox1, which might positively regulate the insertion of the heme cofactor into Cox1. (It is currently debated whether Shy1 and Mss51 exist in the same complex with Cox1, since different experimental approaches lead to opposite results.) Subsequent addition of further, nuclear-encoded subunits of cytochrome oxidase, such as Cox6, leads to the release of Mss51 from the assembly intermediates, which allows further rounds of Cox1 synthesis. The mitochondrial Hsp70 Ssc1, which has been suggested to associate with Mss51 in the ‘TE’ and in the ‘SI’ states is not shown.
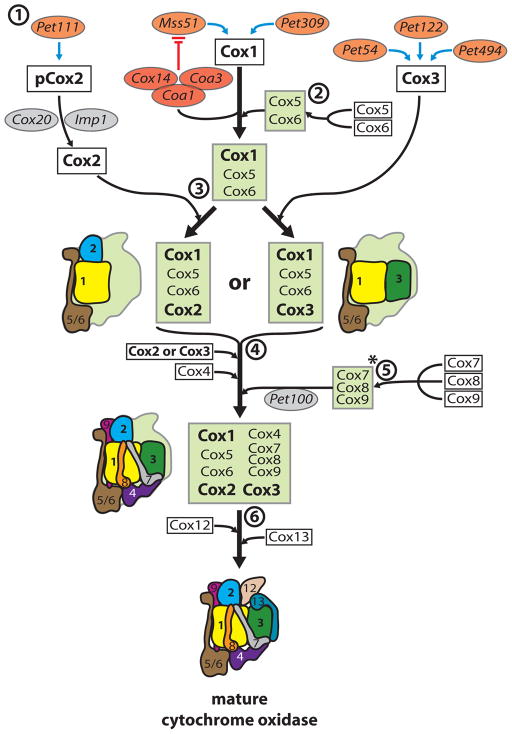
References
-
- Schmidt O, Pfanner N, Meisinger C. Mitochondrial protein import: from proteomics to functional mechanisms. Nat Rev Mol Cell Biol. 2010;11:655–667. - PubMed
-
- Vögtle FN, et al. Global analysis of the mitochondrial N-proteome identifies a processing peptidase critical for protein stability. Cell. 2009;139:428–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
