

S-glutathionylation uncouples eNOS and regulates its cellular and vascular function
- PMID: 21179168
- PMCID: PMC3370391
- DOI: 10.1038/nature09599
S-glutathionylation uncouples eNOS and regulates its cellular and vascular function
Abstract
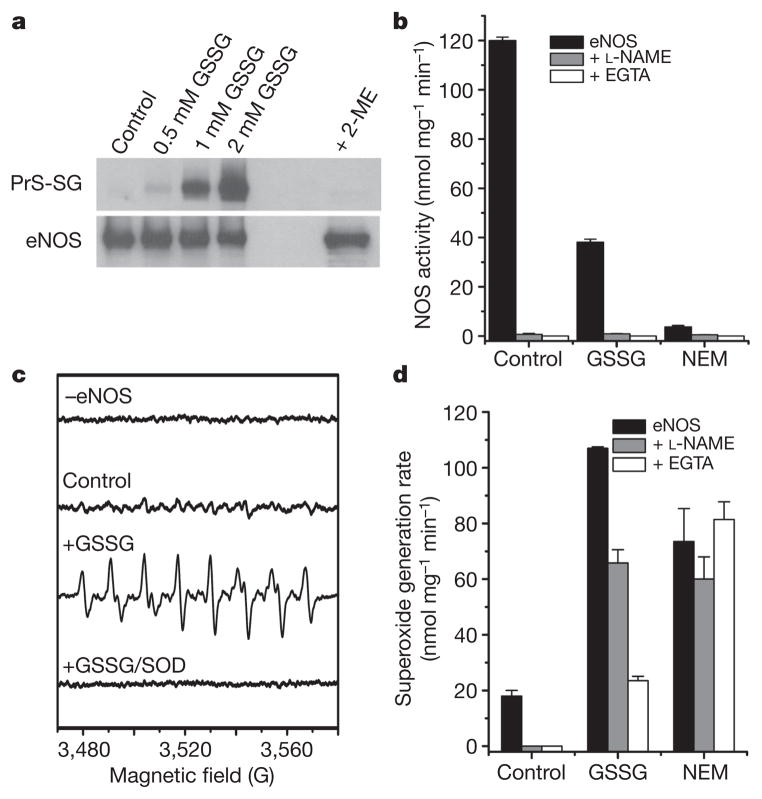
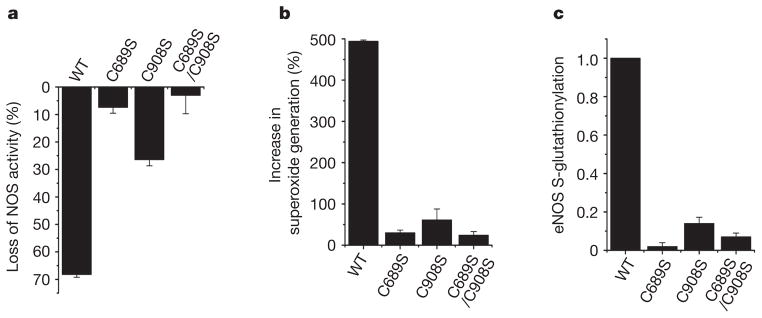
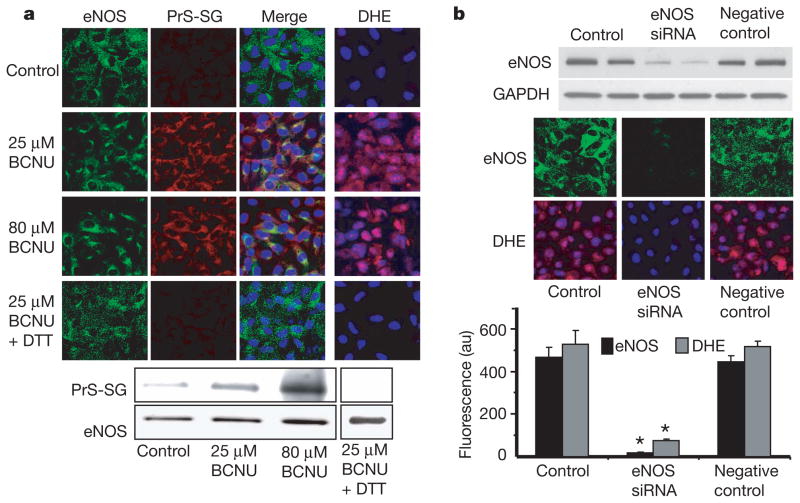
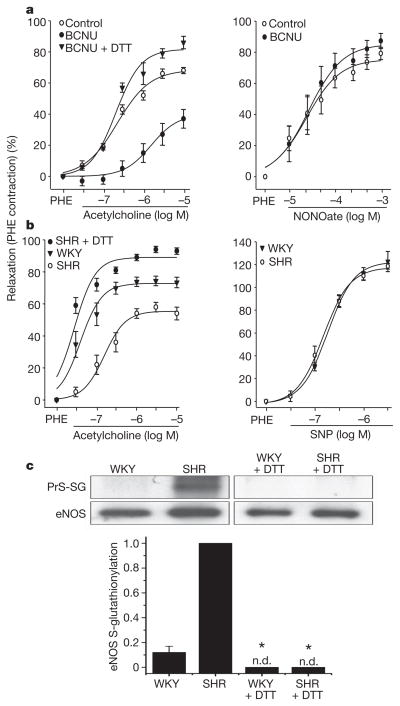
Endothelial nitric oxide synthase (eNOS) is critical in the regulation of vascular function, and can generate both nitric oxide (NO) and superoxide (O(2)(•-)), which are key mediators of cellular signalling. In the presence of Ca(2+)/calmodulin, eNOS produces NO, endothelial-derived relaxing factor, from l-arginine (l-Arg) by means of electron transfer from NADPH through a flavin containing reductase domain to oxygen bound at the haem of an oxygenase domain, which also contains binding sites for tetrahydrobiopterin (BH(4)) and l-Arg. In the absence of BH(4), NO synthesis is abrogated and instead O(2)(•-) is generated. While NOS dysfunction occurs in diseases with redox stress, BH(4) repletion only partly restores NOS activity and NOS-dependent vasodilation. This suggests that there is an as yet unidentified redox-regulated mechanism controlling NOS function. Protein thiols can undergo S-glutathionylation, a reversible protein modification involved in cellular signalling and adaptation. Under oxidative stress, S-glutathionylation occurs through thiol-disulphide exchange with oxidized glutathione or reaction of oxidant-induced protein thiyl radicals with reduced glutathione. Cysteine residues are critical for the maintenance of eNOS function; we therefore speculated that oxidative stress could alter eNOS activity through S-glutathionylation. Here we show that S-glutathionylation of eNOS reversibly decreases NOS activity with an increase in O(2)(•-) generation primarily from the reductase, in which two highly conserved cysteine residues are identified as sites of S-glutathionylation and found to be critical for redox-regulation of eNOS function. We show that eNOS S-glutathionylation in endothelial cells, with loss of NO and gain of O(2)(•-) generation, is associated with impaired endothelium-dependent vasodilation. In hypertensive vessels, eNOS S-glutathionylation is increased with impaired endothelium-dependent vasodilation that is restored by thiol-specific reducing agents, which reverse this S-glutathionylation. Thus, S-glutathionylation of eNOS is a pivotal switch providing redox regulation of cellular signalling, endothelial function and vascular tone.
Figures
References
-
- Bredt DS, et al. Cloned and expressed nitric oxide synthase structurally resembles cytochrome P-450 reductase. Nature. 1991;351:714–718. - PubMed
-
- Palmer RM, Ashton DS, Moncada S. Vascular endothelial cells synthesize nitric oxide from L-arginine. Nature. 1988;333:664–666. - PubMed
-
- Rapoport RM, Draznin MB, Murad F. Endothelium-dependent relaxation in rat aorta may be mediated through cyclic GMP-dependent protein phosphorylation. Nature. 1983;306:174–176. - PubMed
-
- Xia Y, Tsai AL, Berka V, Zweier JL. Superoxide generation from endothelial nitric-oxide synthase. A Ca2+/calmodulin-dependent and tetrahydrobiopterin regulatory process. J Biol Chem. 1998;273:25804–25808. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
