

Engineering the chloroplast targeted malarial vaccine antigens in Chlamydomonas starch granules
- PMID: 21179538
- PMCID: PMC3002285
- DOI: 10.1371/journal.pone.0015424
Engineering the chloroplast targeted malarial vaccine antigens in Chlamydomonas starch granules
Abstract
Background: Malaria, an Anopheles-borne parasitic disease, remains a major global health problem causing illness and death that disproportionately affects developing countries. Despite the incidence of malaria, which remains one of the most severe infections of human populations, there is no licensed vaccine against this life-threatening disease. In this context, we decided to explore the expression of Plasmodium vaccine antigens fused to the granule bound starch synthase (GBSS), the major protein associated to the starch matrix in all starch-accumulating plants and algae such as Chlamydomonas reinhardtii.
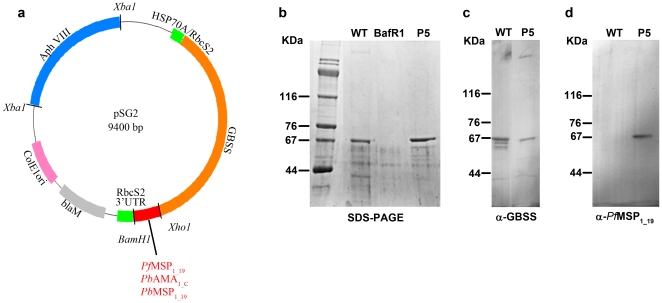
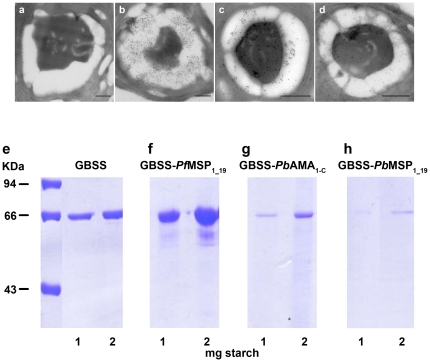
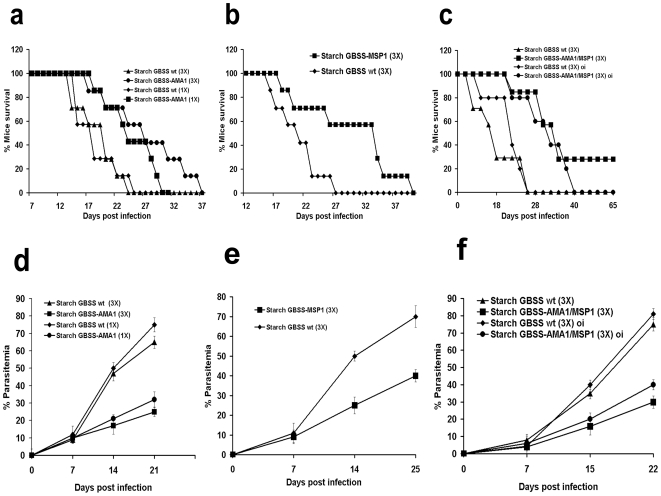
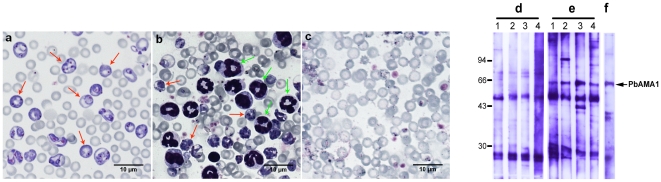
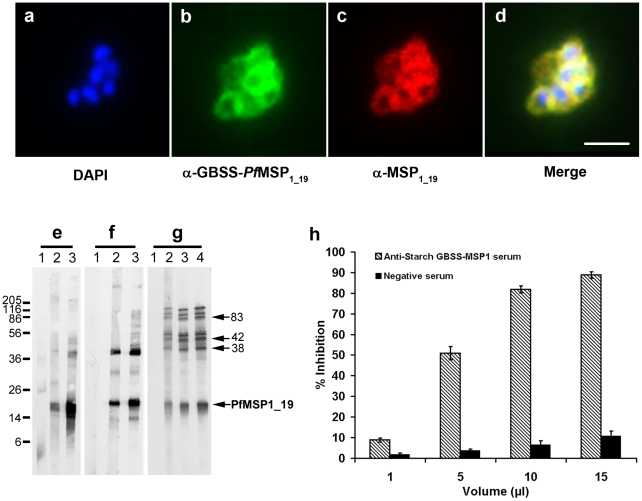
Methods and findings: We describe the development of genetically engineered starch granules containing plasmodial vaccine candidate antigens produced in the unicellular green algae Chlamydomonas reinhardtii. We show that the C-terminal domains of proteins from the rodent Plasmodium species, Plasmodium berghei Apical Major Antigen AMA1, or Major Surface Protein MSP1 fused to the algal granule bound starch synthase (GBSS) are efficiently expressed and bound to the polysaccharide matrix. Mice were either immunized intraperitoneally with the engineered starch particles and Freund adjuvant, or fed with the engineered particles co-delivered with the mucosal adjuvant, and challenged intraperitoneally with a lethal inoculum of P. Berghei. Both experimental strategies led to a significantly reduced parasitemia with an extension of life span including complete cure for intraperitoneal delivery as assessed by negative blood thin smears. In the case of the starch bound P. falciparum GBSS-MSP1 fusion protein, the immune sera or purified immunoglobulin G of mice immunized with the corresponding starch strongly inhibited in vitro the intra-erythrocytic asexual development of the most human deadly plasmodial species.
Conclusion: This novel system paves the way for the production of clinically relevant plasmodial antigens as algal starch-based particles designated herein as amylosomes, demonstrating that efficient production of edible vaccines can be genetically produced in Chlamydomonas.
Conflict of interest statement
Figures
References
-
- WHO, World malaria report, 2008. Available: http://apps.who.int/malaria/wnr2008/malaria2008.pdf.
-
- World Health Organization. Severe falciparum malaria. Trans R Soc Trop Med Hyg. 2000;94:S1–S90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
