

Sorting mouse jejunal epithelial cells with CD24 yields a population with characteristics of intestinal stem cells
- PMID: 21183658
- PMCID: PMC3064119
- DOI: 10.1152/ajpgi.00453.2010
Sorting mouse jejunal epithelial cells with CD24 yields a population with characteristics of intestinal stem cells
Abstract
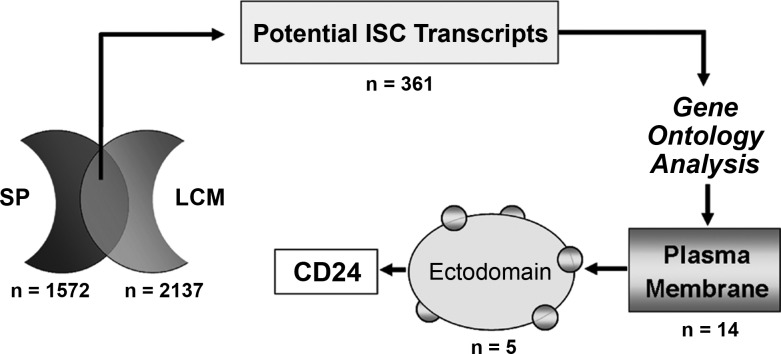
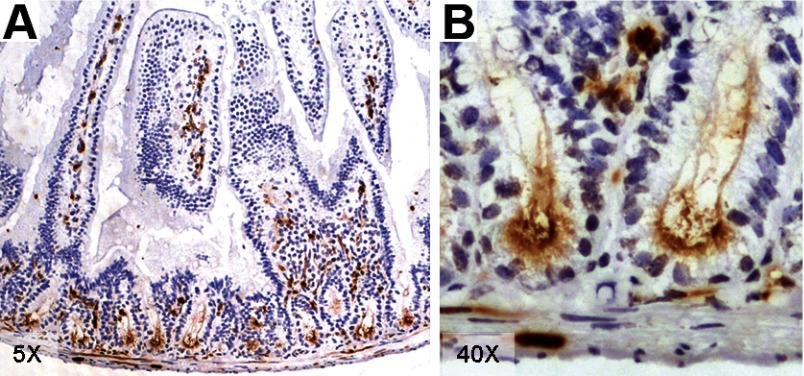
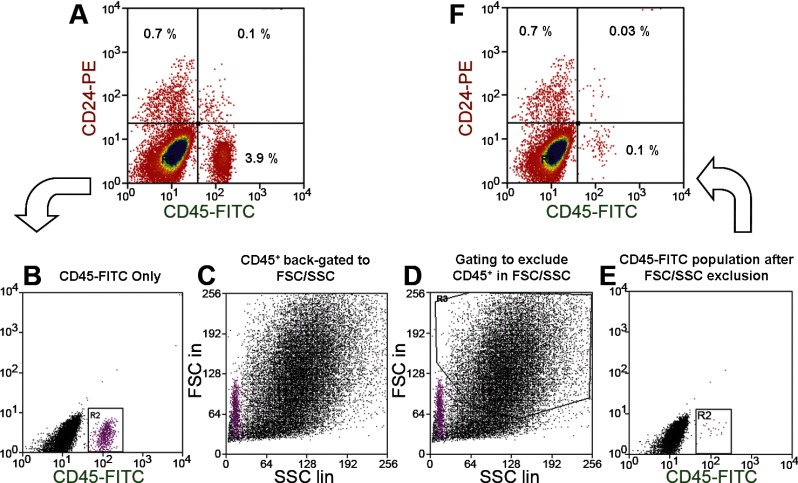
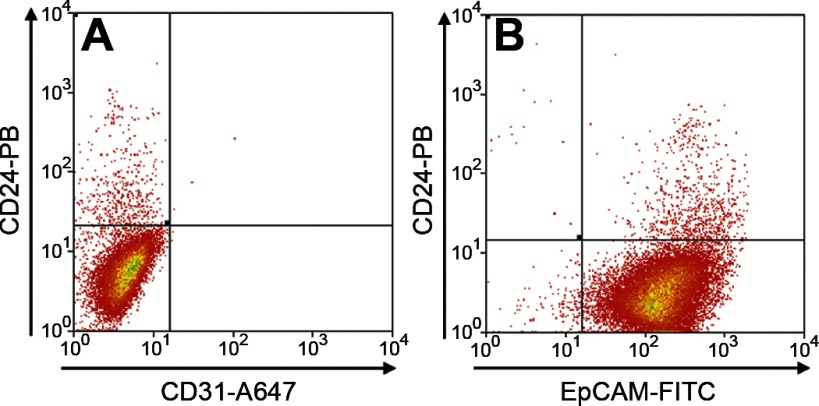
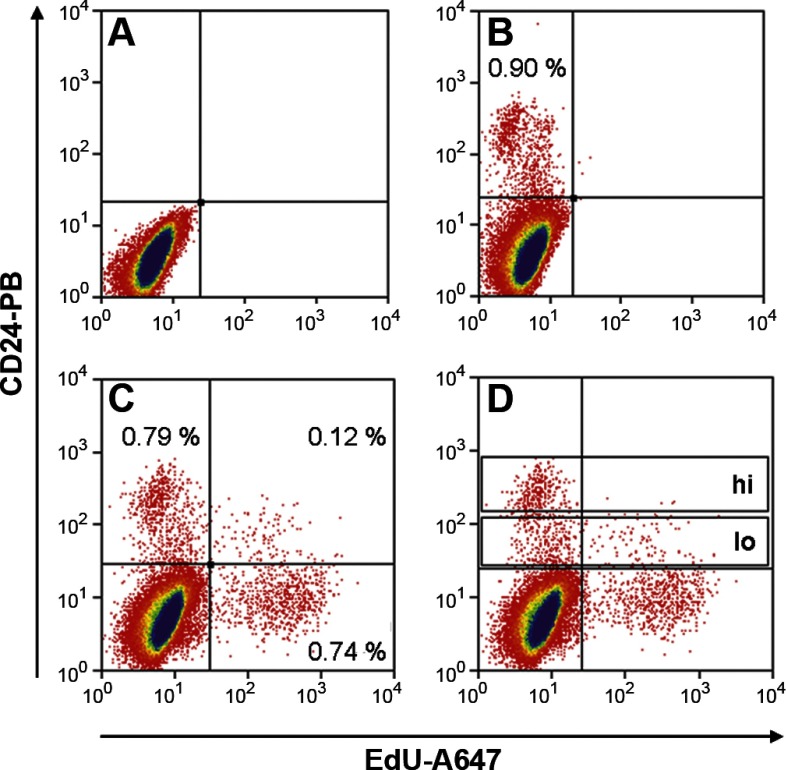
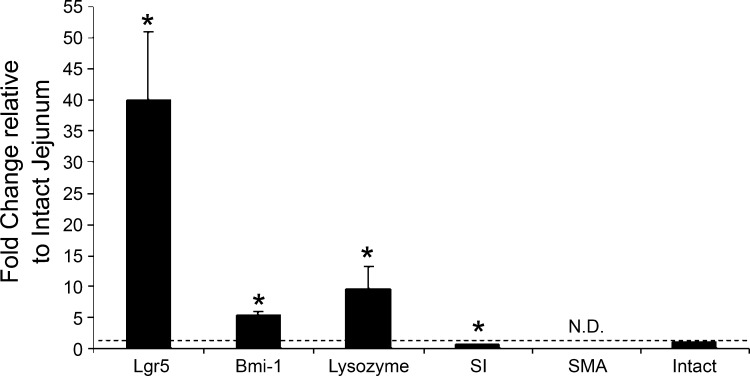
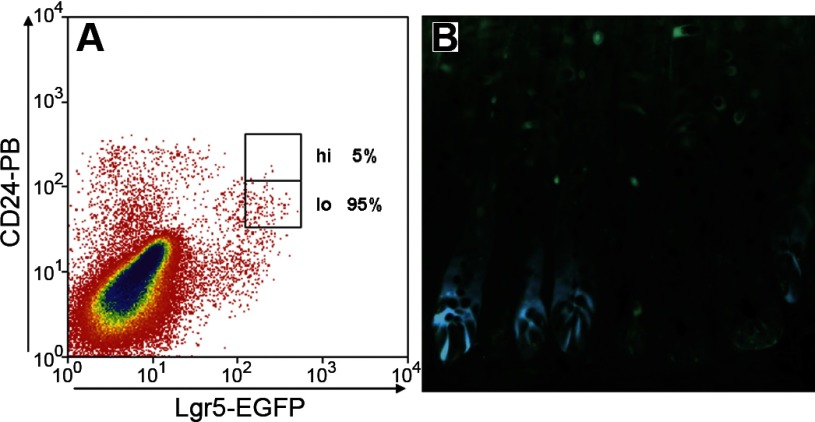
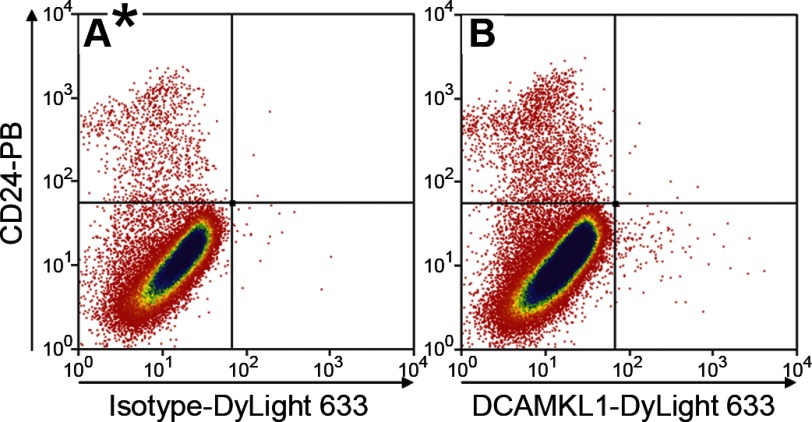
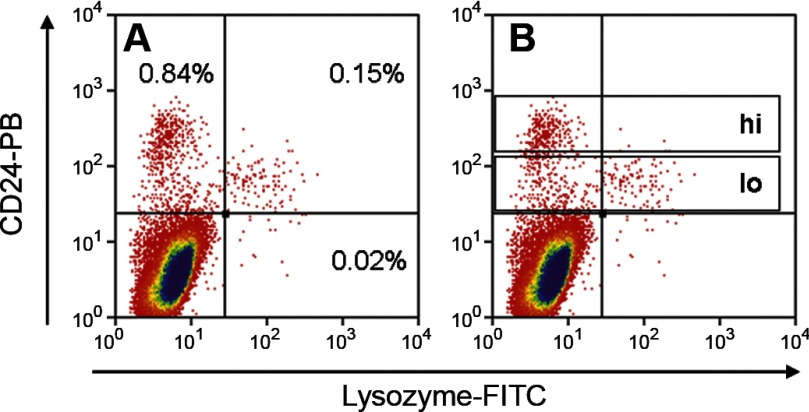
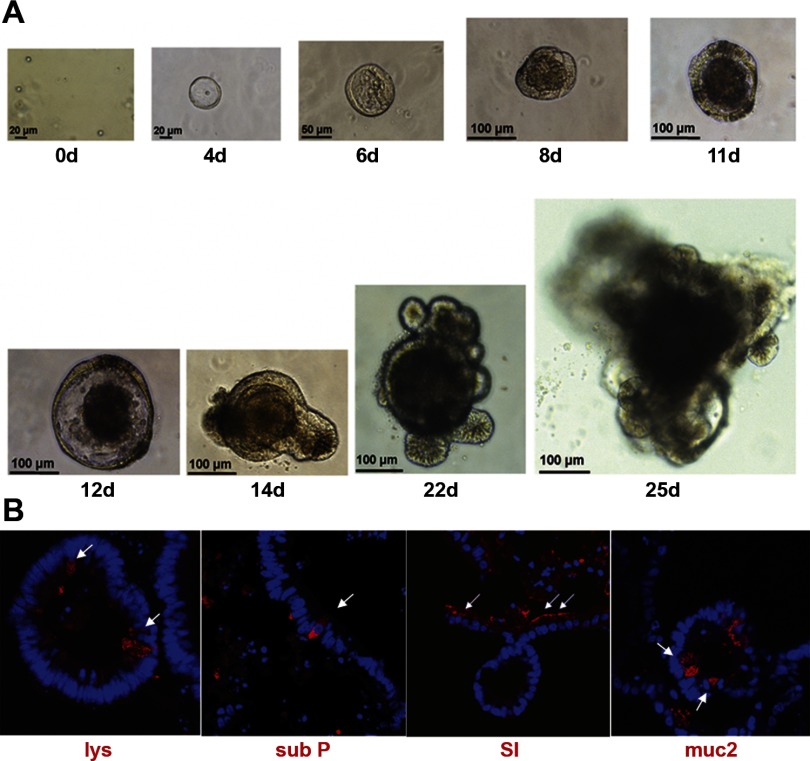
Intestinal stem cells (ISCs) have been studied for more than three decades; however, their isolation has remained a challenge. We hypothesized that, just as for stem cells of other tissues, one or more membrane markers would allow positive selection of ISCs by antibody-based sorting. To explore this hypothesis, microarray data of putative ISC fractions generated by side population sorting and laser capture microdissection were subjected to bioinformatic analysis to identify common membrane antigens. The microarray comparison suggested CD24 as a candidate surface marker, and immunohistochemistry showed expression of CD24 in epithelial cells of crypt bases. Flow cytometry of jejunal epithelial preparations revealed a CD24(+) CD45(-) fraction comprising ∼1% of the cells. Analysis with epithelial cell adhesion molecule and CD31 confirmed that the cell preparations were epithelial and without endothelial contamination. Cycling cells identified by prior injection with 5-ethynyl-2'-deoxyuridine were found predominantly in the CD24(lo) subfraction. Transcript analysis by real-time RT-PCR showed this subfraction to be enriched in the ISC markers leucine-rich-repeat-containing G-protein-coupled receptor 5 (40-fold) and Bmi1 (5-fold), but also enriched in lysozyme (10-fold). Flow cytometry with anti-lysozyme antibodies demonstrated that Paneth cells comprise ∼30% of the CD24(lo) subfraction. Additional flow analyses with leucine-rich-repeat-containing G-protein-coupled receptor 5-enhanced green fluorescent protein (EGFP) epithelium demonstrated colocalization of EGFP(hi) and CD24(lo). In contrast, CD24 cells were negative for the quiescent ISC marker doublecortin and CaM kinase-like-1. Culture of CD24(lo) cells in Matrigel generated organoid structures, which included all four epithelial lineages, thus giving functional evidence for the presence of ISCs. We conclude that the CD24(lo) fraction of jejunal epithelium is highly enriched with cycling ISCs. This isolation method should be useful to many investigators in the field to advance both the basic understanding of ISC biology and the therapeutic applications of ISCs.
Figures
References
-
- Balzar M, Winter MJ, de Boer CJ, Litvinov SV. The biology of the 17–1A antigen (Ep-CAM). J Mol Med 77: 699–712, 1999. - PubMed
-
- Barker N, van Es JH, Kuipers J, Kujala P, van den BM, Cozijnsen M, Haegebarth A, Korving J, Begthel H, Peters PJ, Clevers H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 449: 1003–1007, 2007. - PubMed
-
- Bjerknes M, Cheng H. Intestinal epithelial stem cells and progenitors. Methods Enzymol 419: 337–383, 2006. - PubMed
-
- Cheng H, Leblond CP. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine. V. Unitarian theory of the origin of the four epithelial cell types. Am J Anat 141: 537–562, 1974. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous