

The Atg6/Vps30/Beclin 1 ortholog BEC-1 mediates endocytic retrograde transport in addition to autophagy in C. elegans
- PMID: 21183797
- PMCID: PMC3108013
- DOI: 10.4161/auto.7.4.14391
The Atg6/Vps30/Beclin 1 ortholog BEC-1 mediates endocytic retrograde transport in addition to autophagy in C. elegans
Abstract
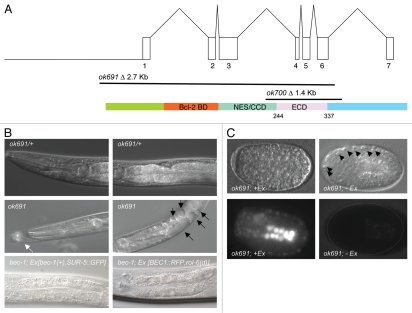
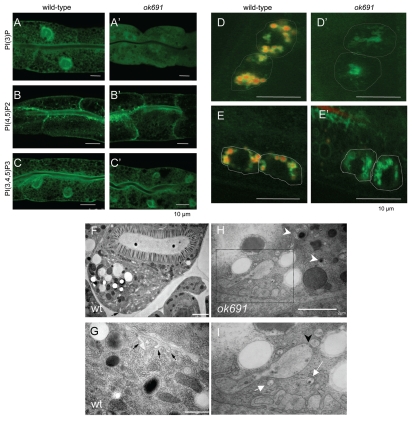
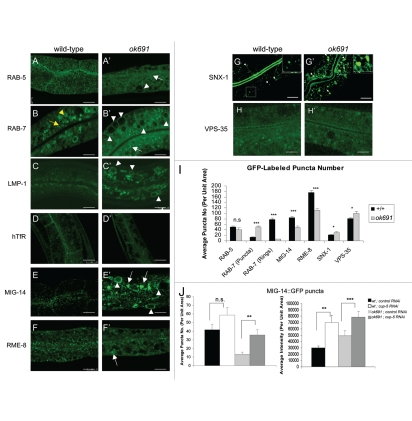
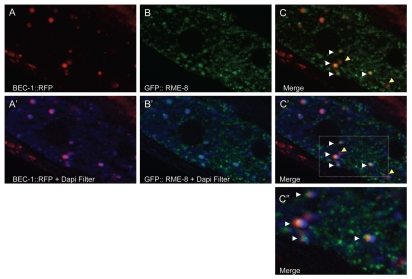
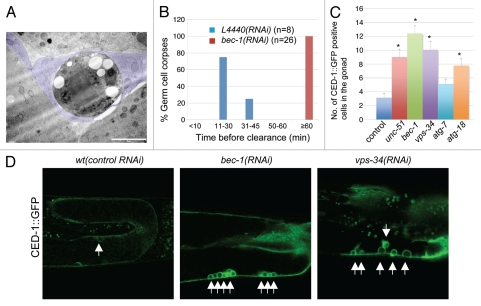
Autophagy and endocytosis are dynamic and tightly regulated processes that contribute to many fundamental aspects of biology including survival, longevity, and development. However, the molecular links between autophagy and endocytosis are not well understood. Here, we report that BEC-1, the C. elegans ortholog of Atg6/Vps30/Beclin1, a key regulator of the autophagic machinery, also contributes to endosome function. In particular we identify a defect in retrograde transport from endosomes to the Golgi in bec-1 mutants. MIG-14/Wntless is normally recycled from endosomes to the Golgi through the action of the retromer complex and its associated factor RME-8. Lack of retromer or RME-8 activity results in the aberrant transport of MIG-14/Wntless to the lysosome where it is degraded. Similarly, we find that lack of bec-1 also results in mislocalization and degradation of MIG-14::GFP, reduced levels of RME-8 on endosomal membranes, and the accumulation of morphologically abnormal endosomes. A similar phenotype was observed in animals treated with dsRNA against vps-34. We further identify a requirement for BEC-1 in the clearance of apoptotic corpses in the hermaphrodite gonad, suggesting a role for BEC-1 in phagosome maturation, a process that appears to depend upon retrograde transport. In addition, autophagy genes may also be required for cell corpse clearance, as we find that RNAi against atg-18 or unc-51 also results in a lack of cell corpse clearance.
Figures
References
-
- Suzuki K, Kubota Y, Sekito T, Ohsumi Y. Hierarchy of Atg proteins in pre-autophagosomal structure organization. Genes Cells. 2007;12:209–218. - PubMed
-
- Klionsky DJ, Cregg JM, Dunn W, Jr, Emr SD, Sakai Y, Sandoval IV, et al. A unified nomenclature for yeast autophagy-related genes. Dev Cell. 2003;5:539–545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials