

Molecular control of polyene macrolide biosynthesis: direct binding of the regulator PimM to eight promoters of pimaricin genes and identification of binding boxes
- PMID: 21187288
- PMCID: PMC3059063
- DOI: 10.1074/jbc.M110.182428
Molecular control of polyene macrolide biosynthesis: direct binding of the regulator PimM to eight promoters of pimaricin genes and identification of binding boxes
Abstract
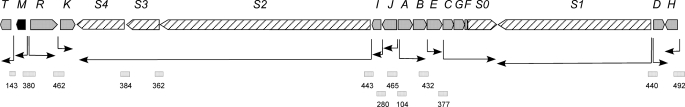
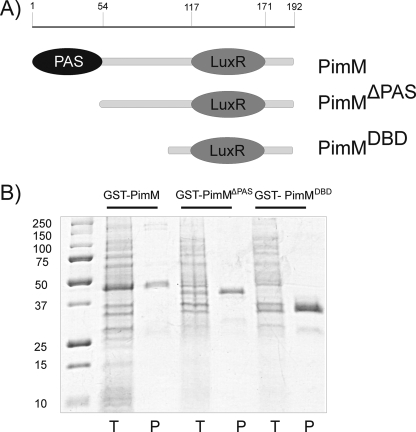
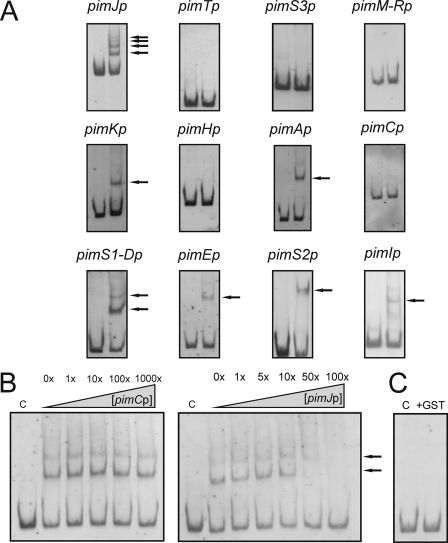
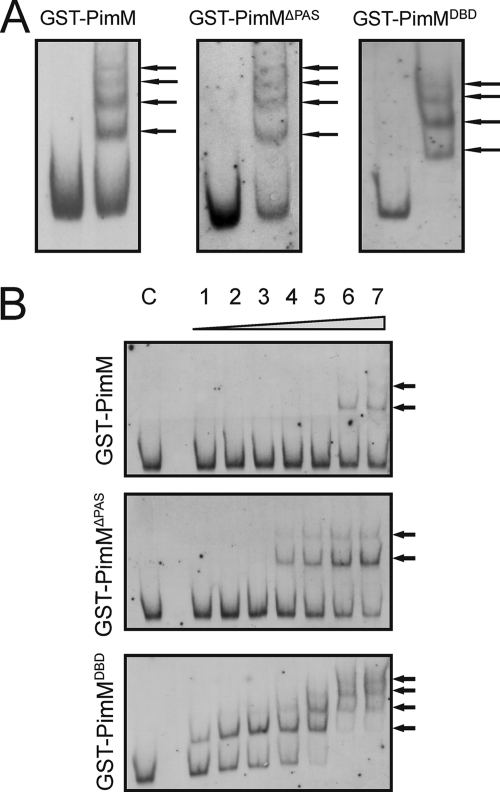
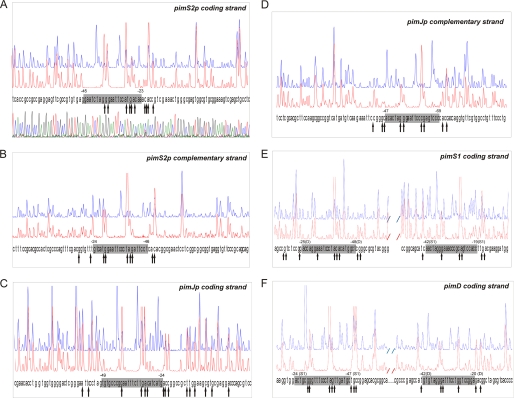
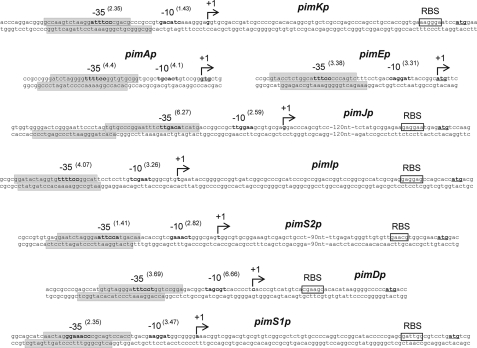
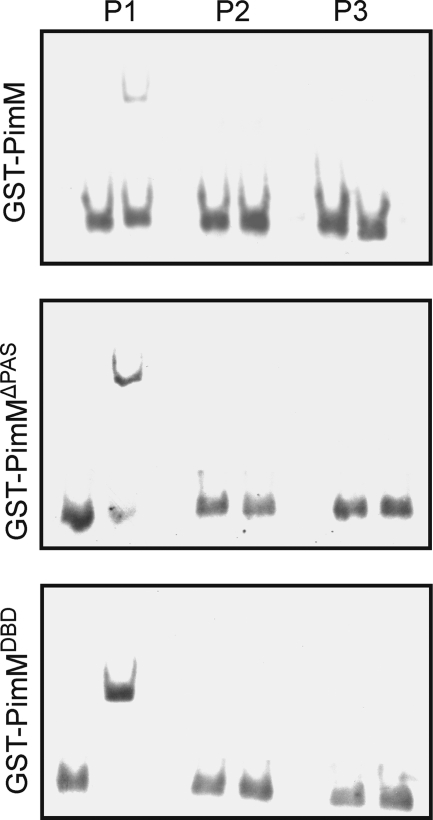
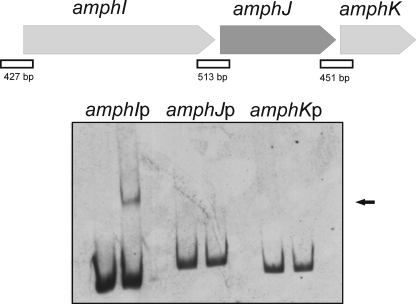
Control of polyene macrolide production in Streptomyces natalensis is mediated by the transcriptional activator PimM. This regulator, which combines an N-terminal PAS domain with a C-terminal helix-turn-helix motif, is highly conserved among polyene biosynthetic gene clusters. PimM, truncated forms of the protein without the PAS domain (PimM(ΔPAS)), and forms containing just the DNA-binding domain (DBD) (PimM(DBD)) were overexpressed in Escherichia coli as GST-fused proteins. GST-PimM binds directly to eight promoters of the pimaricin cluster, as demonstrated by electrophoretic mobility shift assays. Assays with truncated forms of the protein revealed that the PAS domain does not mediate specificity or the distinct recognition of target genes, which rely on the DBD domain, but significantly reduces binding affinity up to 500-fold. Transcription start points were identified by 5'-rapid amplification of cDNA ends, and the binding regions of PimM(DBD) were investigated by DNase I protection studies. In all cases, binding took place covering the -35 hexamer box of each promoter, suggesting an interaction of PimM and RNA polymerase to cause transcription activation. Information content analysis of the 16 sequences protected in target promoters was used to deduce the structure of the PimM-binding site. This site displays dyad symmetry, spans 14 nucleotides, and adjusts to the consensus TVGGGAWWTCCCBA. Experimental validation of this binding site was performed by using synthetic DNA duplexes. Binding of PimM to the promoter region of one of the polyketide synthase genes from the Streptomyces nodosus amphotericin cluster containing the consensus binding site was also observed, thus proving the applicability of the findings reported here to other antifungal polyketides.
Figures

References
-
- Bibb M. (1996) Microbiology 142, 1335–1344 - PubMed
-
- Aparicio J. F., Mendes M. V., Antón N., Recio E., Martín J. F. (2004) Curr. Med. Chem. 11, 1645–1656 - PubMed
-
- te Welscher Y. M., ten Napel H. H., Balagué M. M., Souza C. M., Riezman H., de Kruijff B., Breukink E. (2008) J. Biol. Chem. 283, 6393–6401 - PubMed
-
- Caffrey P., Aparicio J. F., Malpartida F., Zotchev S. B. (2008) Curr. Top. Med. Chem. 8, 639–653 - PubMed
-
- Mendes M. V., Recio E., Fouces R., Luiten R., Martín J. F., Aparicio J. F. (2001) Chem. Biol. 8, 635–644 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
