

Biglycan recruits utrophin to the sarcolemma and counters dystrophic pathology in mdx mice
- PMID: 21187385
- PMCID: PMC3021068
- DOI: 10.1073/pnas.1013067108
Biglycan recruits utrophin to the sarcolemma and counters dystrophic pathology in mdx mice
Abstract
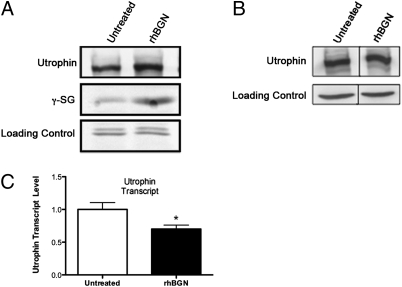
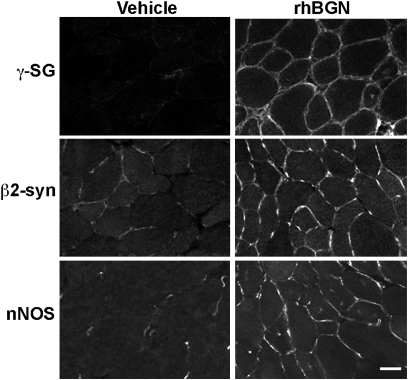
Duchenne muscular dystrophy (DMD) is caused by mutations in dystrophin and the subsequent disruption of the dystrophin-associated protein complex (DAPC). Utrophin is a dystrophin homolog expressed at high levels in developing muscle that is an attractive target for DMD therapy. Here we show that the extracellular matrix protein biglycan regulates utrophin expression in immature muscle and that recombinant human biglycan (rhBGN) increases utrophin expression in cultured myotubes. Systemically delivered rhBGN up-regulates utrophin at the sarcolemma and reduces muscle pathology in the mdx mouse model of DMD. RhBGN treatment also improves muscle function as judged by reduced susceptibility to eccentric contraction-induced injury. Utrophin is required for the rhBGN therapeutic effect. Several lines of evidence indicate that biglycan acts by recruiting utrophin protein to the muscle membrane. RhBGN is well tolerated in animals dosed for as long as 3 months. We propose that rhBGN could be a therapy for DMD.
Conflict of interest statement
Conflict of interest statement: A.R.A., B.A.M., and J.R.F. are inventors on patents owned by Brown University covering biglycan as a therapeutic for neuromuscular diseases. J.F. is a Director and shareholder in Tivorsan pharmaceuticals.
Figures




References
-
- Emery AEH. Duchenne Muscular Dystrophy. 2nd Ed. Oxford: Oxford University Press; 1993.
-
- Koenig M, et al. Complete cloning of the Duchenne muscular dystrophy (DMD) cDNA and preliminary genomic organization of the DMD gene in normal and affected individuals. Cell. 1987;50:509–517. - PubMed
-
- Blake DJ, Weir A, Newey SE, Davies KE. Function and genetics of dystrophin and dystrophin-related proteins in muscle. Physiol Rev. 2002;82:291–329. - PubMed
-
- Muntoni F, Torelli S, Ferlini A. Dystrophin and mutations: One gene, several proteins, multiple phenotypes. Lancet Neurol. 2003;2:731–740. - PubMed
-
- Tinsley J, et al. Expression of full-length utrophin prevents muscular dystrophy in mdx mice. Nat Med. 1998;4:1441–1444. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- KO8 HL072332/HL/NHLBI NIH HHS/United States
- RR15578/RR/NCRR NIH HHS/United States
- T32 MH020068/MH/NIMH NIH HHS/United States
- K08 HL072332/HL/NHLBI NIH HHS/United States
- AR57698/AR/NIAMS NIH HHS/United States
- R01 HD023924/HD/NICHD NIH HHS/United States
- R01 EY013862/EY/NEI NIH HHS/United States
- P20 RR018757/RR/NCRR NIH HHS/United States
- EY 013862/EY/NEI NIH HHS/United States
- R01 AR048871/AR/NIAMS NIH HHS/United States
- AR 48871/AR/NIAMS NIH HHS/United States
- R21 AR055878/AR/NIAMS NIH HHS/United States
- P20 RR015578/RR/NCRR NIH HHS/United States
- HD23924/HD/NICHD NIH HHS/United States
- NS064295/NS/NINDS NIH HHS/United States
- U01 NS064295/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
