

Arc-dependent synapse-specific homeostatic plasticity
- PMID: 21187403
- PMCID: PMC3021034
- DOI: 10.1073/pnas.1017914108
Arc-dependent synapse-specific homeostatic plasticity
Abstract
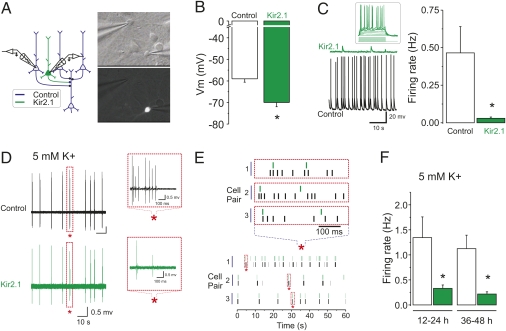
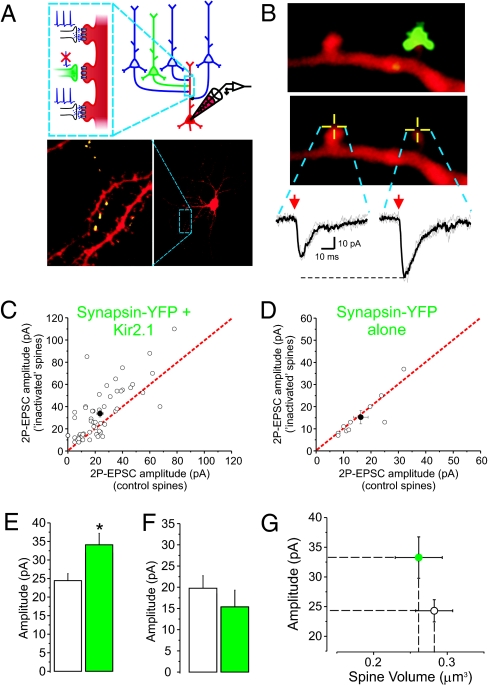
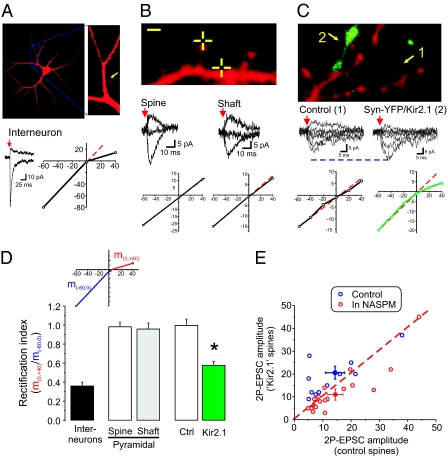
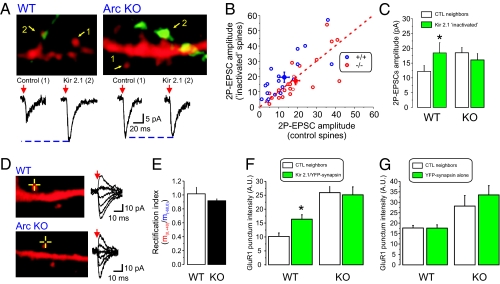
Both theoretical and experimental research has indicated that the synaptic strength between neurons in a network needs to be properly fine-tuned and controlled by homeostatic mechanisms to ensure proper network function. One such mechanism that has been extensively characterized is synaptic homeostatic plasticity or global synaptic scaling. This mechanism refers to the bidirectional ability of all synapses impinging on a neuron to actively compensate for changes in the neuron's overall excitability. Here, using a combination of electrophysiological, two-photon glutamate uncaging and imaging methods, we show that mature individual synapses, independent of neighboring synapses, have the ability to autonomously sense their level of activity and actively compensate for it in a homeostatic-like fashion. This synapse-specific homeostatic plasticity, similar to global synaptic plasticity, requires the immediate early gene Arc. Together, our results document an extra level of regulation of synaptic function that bears important computational consequences on information storage in the brain.
Conflict of interest statement
Conflict of interest statement: Under a licensing agreement between Millipore Corporation and The Johns Hopkins University, R.L.H. is entitled to a share of royalties received by the University on sales of products described in this article. R.L.H. is a paid consultant to Millipore Corporation. The terms of this arrangement are managed by The Johns Hopkins University in accordance with its conflict of interest policies.
Figures
References
-
- Abbott LF, Nelson SB. Synaptic plasticity: Taming the beast. Nat Neurosci. 2000;3(Suppl):1178–1183. - PubMed
-
- O'Brien RJ, et al. Activity-dependent modulation of synaptic AMPA receptor accumulation. Neuron. 1998;21:1067–1078. - PubMed
-
- Turrigiano GG, Leslie KR, Desai NS, Rutherford LC, Nelson SB. Activity-dependent scaling of quantal amplitude in neocortical neurons. Nature. 1998;391:892–896. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
