

Structural studies of ion permeation and Ca2+ blockage of a bacterial channel mimicking the cyclic nucleotide-gated channel pore
- PMID: 21187429
- PMCID: PMC3021057
- DOI: 10.1073/pnas.1013643108
Structural studies of ion permeation and Ca2+ blockage of a bacterial channel mimicking the cyclic nucleotide-gated channel pore
Abstract
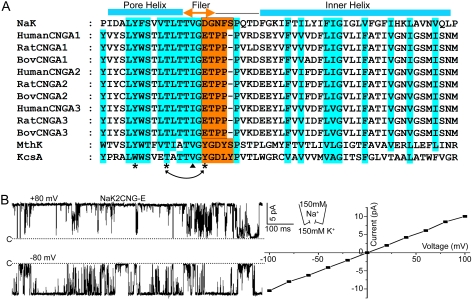
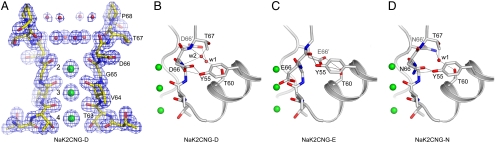
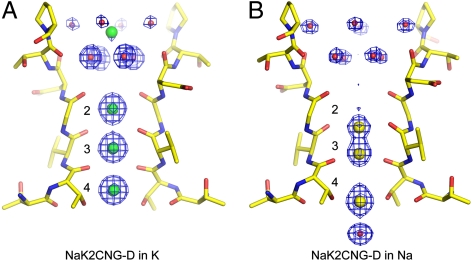
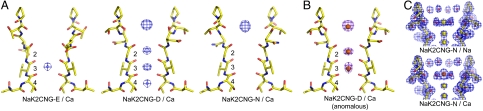
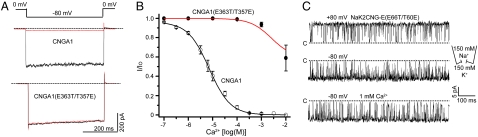
Cyclic nucleotide-gated (CNG) channels play an essential role in the visual and olfactory sensory systems and are ubiquitous in eukaryotes. Details of their underlying ion selectivity properties are still not fully understood and are a matter of debate in the absence of high-resolution structures. To reveal the structural mechanism of ion selectivity in CNG channels, particularly their Ca(2+) blockage property, we engineered a set of mimics of CNG channel pores for both structural and functional analysis. The mimics faithfully represent the CNG channels they are modeled after, permeate Na(+) and K(+) equally well, and exhibit the same Ca(2+) blockage and permeation properties. Their high-resolution structures reveal a hitherto unseen selectivity filter architecture comprising three contiguous ion binding sites in which Na(+) and K(+) bind with different ion-ligand geometries. Our structural analysis reveals that the conserved acidic residue in the filter is essential for Ca(2+) binding but not through direct ion chelation as in the currently accepted view. Furthermore, structural insight from our CNG mimics allows us to pinpoint equivalent interactions in CNG channels through structure-based mutagenesis that have previously not been predicted using NaK or K(+) channel models.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Mechanism of K+/Na+ selectivity in potassium channels from the perspective of the non-selective bacterial channel NaK.Channels (Austin). 2011 May-Jun;5(3):198-200. doi: 10.4161/chan.5.3.15873. Epub 2011 May 1. Channels (Austin). 2011. PMID: 21540644
References
-
- Yau KW, Baylor DA. Cyclic GMP-activated conductance of retinal photoreceptor cells. Annu Rev Neurosci. 1989;12:289–327. - PubMed
-
- Kaupp UB, Seifert R. Cyclic nucleotide-gated ion channels. Physiol Rev. 2002;82:769–824. - PubMed
-
- Matulef K, Zagotta WN. Cyclic nucleotide-gated ion channels. Annu Rev Cell Dev Biol. 2003;19:23–44. - PubMed
-
- Zagotta WN, Siegelbaum SA. Structure and function of cyclic nucleotide-gated channels. Annu Rev Neurosci. 1996;19:235–263. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
