

Immature T-cell clustering and efficient differentiation require the polarity protein Scribble
- PMID: 21189299
- PMCID: PMC3024664
- DOI: 10.1073/pnas.1018224108
Immature T-cell clustering and efficient differentiation require the polarity protein Scribble
Abstract
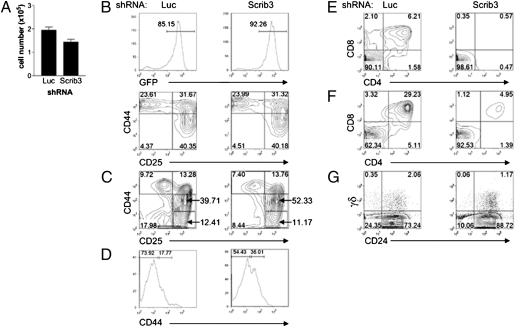
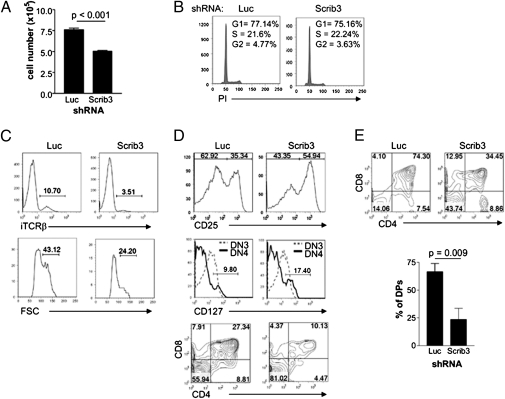
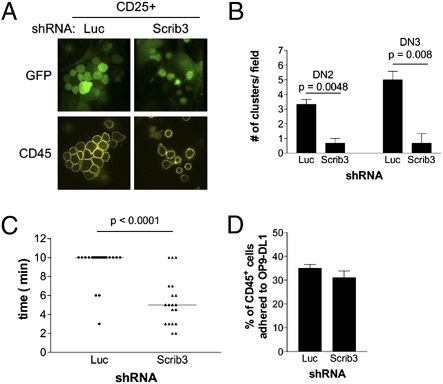
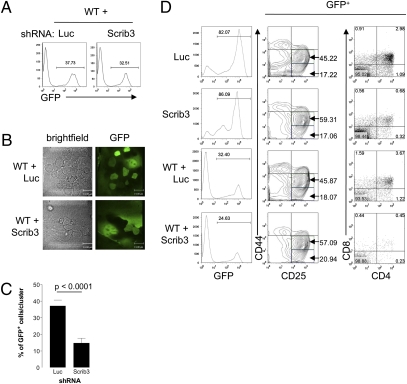
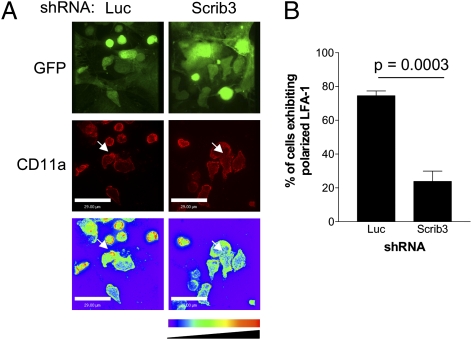
T-cell polarization is required for cell migration and cell-cell interactions, cellular behaviors crucial for lymphocyte differentiation. Despite expression of the epithelial polarity network in T cells, neither its contribution to thymocyte polarity nor its requirement during development is known. We report here that depletion of the polarity protein Scribble in hematopoietic progenitor cells results in inefficient T-cell development characterized by a partial developmental block during the early double-negative (DN) stage of differentiation. Scribble-depleted hematopoietic progenitor cells exhibit a delayed transition into late CD44(lo/-)CD25(+) DN3 cells, evidenced by the accumulation of early CD44(int)CD25(+) DN3 cells. As a consequence, a limited cellular expansion and a reduced frequency of intracellular T-cell receptor β-positive DN3 cells are observed among Scribble-deficient differentiating T cells. Moreover, whereas purified Scribble-depleted DN2 and DN3 cells do not exhibit compromised spontaneous motility, T-cell clustering and prolonged homotypic interactions among such cells are reduced. This deficiency correlates with a lack of polarization of the integrin LFA-1 during T-cell migration or on the initiation of T-cell-T-cell interactions. Scribble is therefore a critical contributor to the clustering of immature T cells, an event shown here to be necessary for efficient developmental progression.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Dow LE, Humbert PO. Polarity regulators and the control of epithelial architecture, cell migration, and tumorigenesis. Int Rev Cytol. 2007;262:253–302. - PubMed
-
- Ladi E, Yin X, Chtanova T, Robey EA. Thymic microenvironments for T cell differentiation and selection. Nat Immunol. 2006;7:338–343. - PubMed
-
- Takahama Y. Journey through the thymus: Stromal guides for T-cell development and selection. Nat Rev Immunol. 2006;6:127–135. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
