

Binding of the inhibitor protein IF(1) to bovine F(1)-ATPase
- PMID: 21192948
- PMCID: PMC3041923
- DOI: 10.1016/j.jmb.2010.12.025
Binding of the inhibitor protein IF(1) to bovine F(1)-ATPase
Abstract
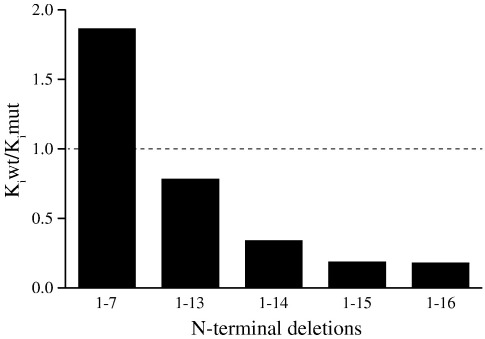
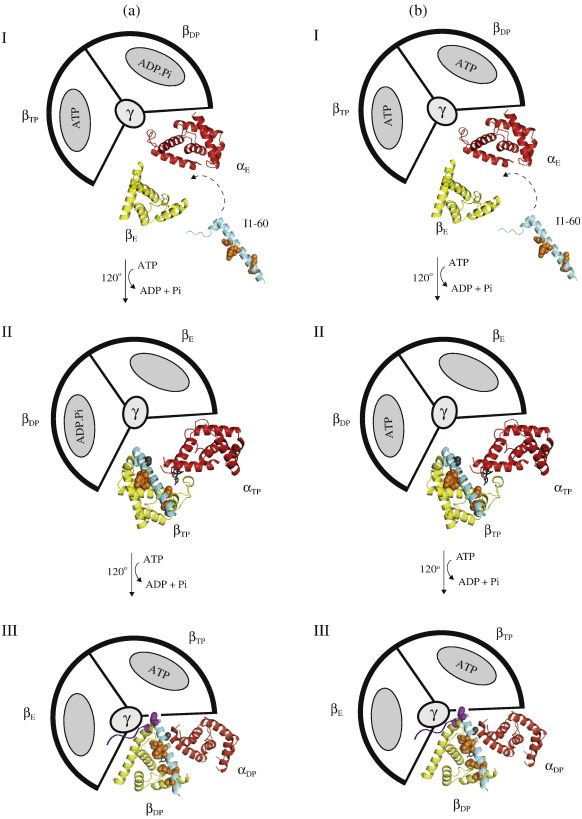
In the structure of bovine F(1)-ATPase inhibited with residues 1-60 of the bovine inhibitor protein IF(1), the α-helical inhibitor interacts with five of the nine subunits of F(1)-ATPase. In order to understand the contributions of individual amino acid residues to this complex binding mode, N-terminal deletions and point mutations have been introduced, and the binding properties of each mutant inhibitor protein have been examined. The N-terminal region of IF(1) destabilizes the interaction of the inhibitor with F(1)-ATPase and may assist in removing the inhibitor from its binding site when F(1)F(o)-ATPase is making ATP. Binding energy is provided by hydrophobic interactions between residues in the long α-helix of IF(1) and the C-terminal domains of the β(DP)-subunit and β(TP)-subunit and a salt bridge between residue E30 in the inhibitor and residue R408 in the C-terminal domain of the β(DP)-subunit. Several conserved charged amino acids in the long α-helix of IF(1) are also required for establishing inhibitory activity, but in the final inhibited state, they are not in contact with F(1)-ATPase and occupy aqueous cavities in F(1)-ATPase. They probably participate in the pathway from the initial interaction of the inhibitor and the enzyme to the final inhibited complex observed in the structure, in which two molecules of ATP are hydrolysed and the rotor of the enzyme turns through two 120° steps. These findings contribute to the fundamental understanding of how the inhibitor functions and to the design of new inhibitors for the systematic analysis of the catalytic cycle of the enzyme.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures






References
-
- Pullman M.E., Monroy G.C. A soluble heat stable protein in mitochondria from bovine heart that inhibits ATP hydrolase activity. J. Biol. Chem. 1963;238:3762–3769. - PubMed
-
- Cabezón E., Montgomery M.G., Leslie A.G.W., Walker J.E. The structure of bovine F1-ATPase in complex with its regulatory protein IF1. Nat. Struct. Biol. 2003;10:744–750. - PubMed
-
- Lippe G., Sorgato M.C., Harris D.A. The binding and release of the inhibitor protein are governed independently by ATP and membrane potential in ox-heart submitochondrial vesicles. Biochim. Biophys. Acta. 1988;933:12–21. - PubMed
-
- Walker J.E., Gay N.J., Powell S.J., Kostina M., Dyer M.R. ATP synthase from bovine mitochondria: sequences of imported precursors of oligomycin sensitivity conferral protein, factor 6, and adenosine triphosphatase inhibitor protein. Biochemistry. 1987;26:8613–8619. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
