

Pulmonary proteases in the cystic fibrosis lung induce interleukin 8 expression from bronchial epithelial cells via a heme/meprin/epidermal growth factor receptor/Toll-like receptor pathway
- PMID: 21193404
- PMCID: PMC3045023
- DOI: 10.1074/jbc.M110.183863
Pulmonary proteases in the cystic fibrosis lung induce interleukin 8 expression from bronchial epithelial cells via a heme/meprin/epidermal growth factor receptor/Toll-like receptor pathway
Abstract
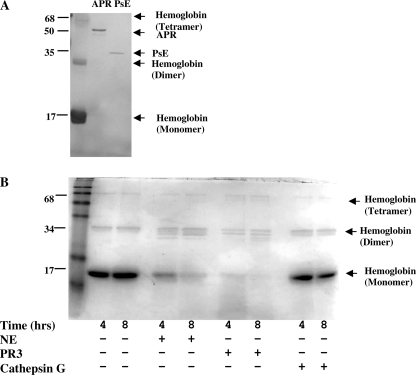
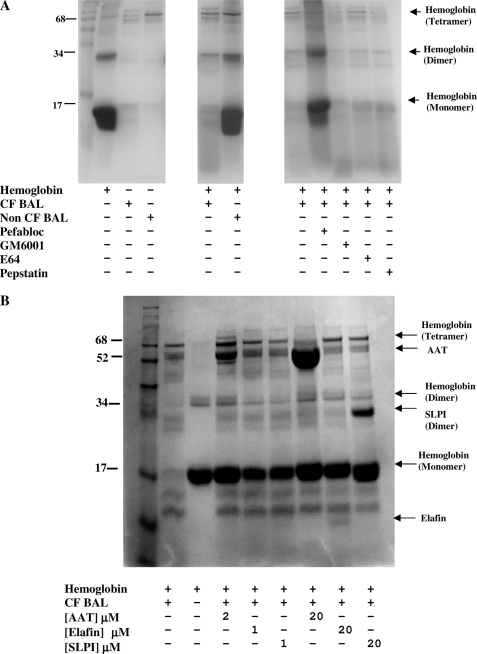
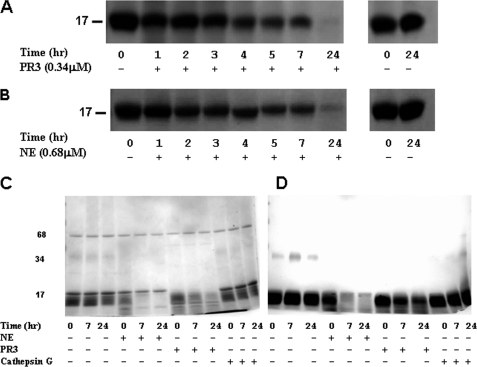
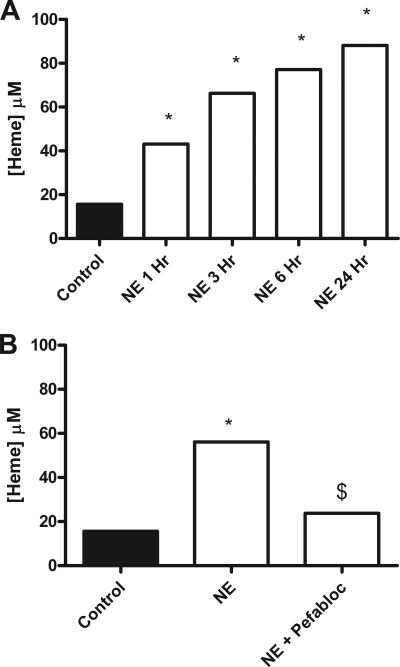
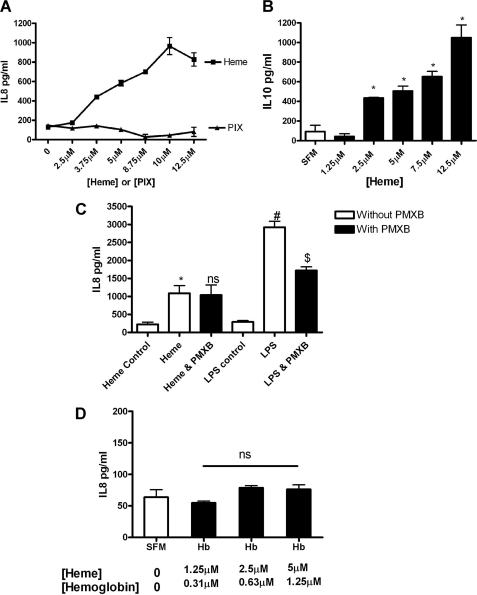
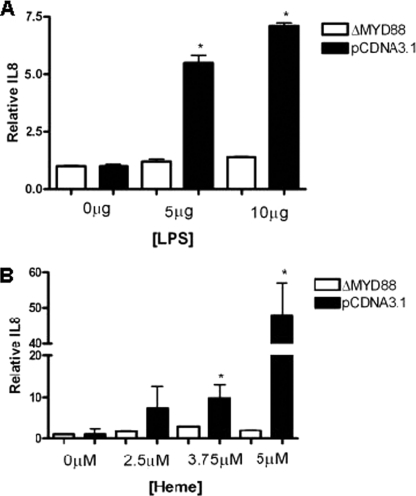
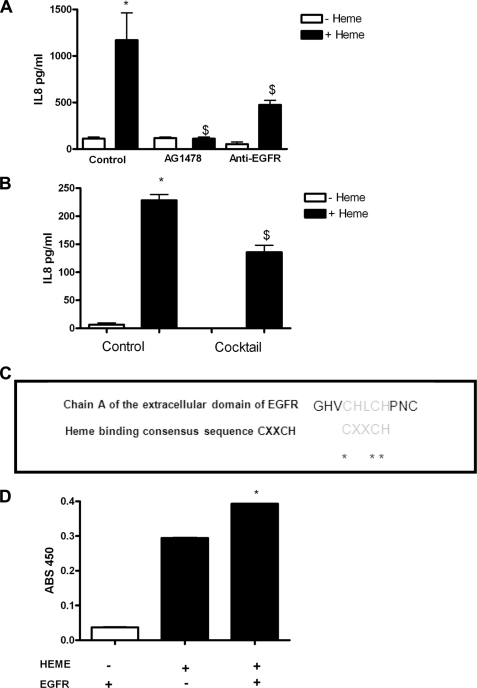
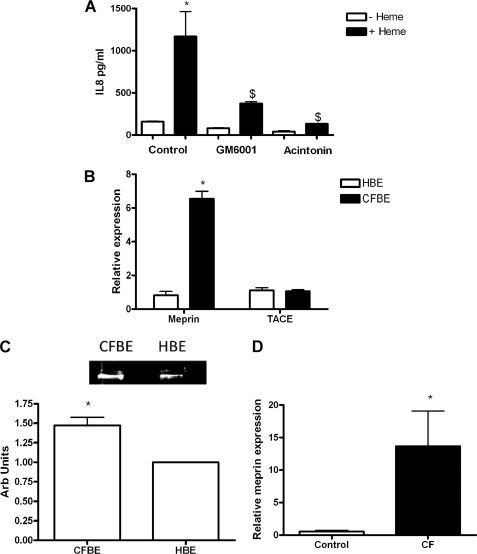
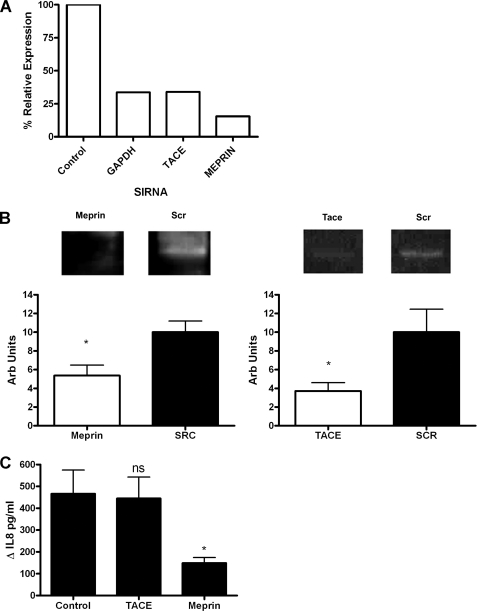
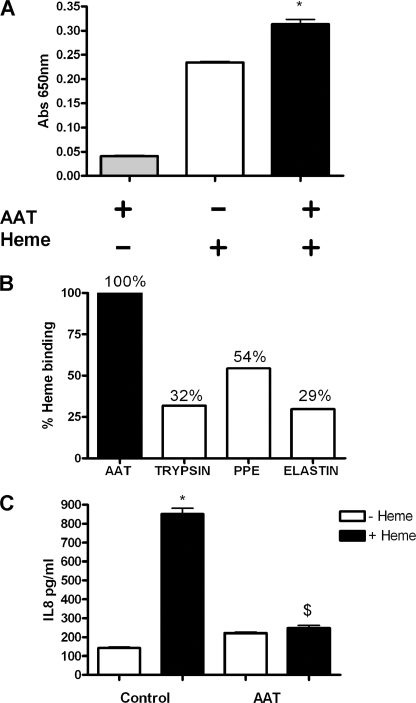
A high intrapulmonary protease burden is characteristic of cystic fibrosis (CF), and the resulting dysregulation of the protease/anti-protease balance has serious implications for inflammation in the CF lung. Because of this inflammation, micro-bleeds can occur releasing hemoglobin into the lung. The aim of this study was to investigate the effect of the protease-rich environment of the CF lung on human hemoglobin and to assess the proinflammatory effect of heme on CF bronchial epithelium. Here, we show that the Pseudomonas proteases (Pseudomonas elastase and alkaline protease) and the neutrophil proteases (neutrophil elastase (NE) and proteinase-3) are capable of almost complete degradation of hemoglobin in vitro but that NE is the predominant protease that cleaves hemoglobin in vivo in CF bronchoalveolar lavage fluid. One of the effects of this is the release of heme, and in this study we show that heme stimulates IL-8 and IL-10 protein production from ΔF508 CFBE41o(-) bronchial epithelial cells. In addition, heme-induced IL-8 expression utilizes a novel pathway involving meprin, EGF receptor, and MyD88. Meprin levels are elevated in CF cell lines and bronchial brushings, thus adding to the proinflammatory milieu. Interestingly, α(1)-antitrypsin, in addition to its ability to neutralize NE and protease-3, can also bind heme and neutralize heme-induced IL-8 from CFBE41o(-) cells. This study illustrates the proinflammatory effects of micro-bleeds in the CF lung, the process by which this occurs, and a potential therapeutic intervention.
Figures
References
-
- Rees D. D., Brain J. D., Wohl M. E., Humes J. L., Mumford R. A. (1997) J. Pharmacol. Exp. Ther. 283, 1201–1206 - PubMed
-
- Geraghty P., Rogan M. P., Greene C. M., Boxio R. M., Poiriert T., O'Mahony M., Belaaouaj A., O'Neill S. J., Taggart C. C., McElvaney N. G. (2007) J. Immunol. 178, 5871–5878 - PubMed
-
- Bergin D. A., Greene C. M., Sterchi E. E., Kenna C., Geraghty P., Belaaouaj A., Belaaouaj A., Taggart C. C., O'Neill S. J., McElvaney N. G. (2008) J. Biol. Chem. 283, 31736–31744 - PubMed
-
- Walsh D. E., Greene C. M., Carroll T. P., Taggart C. C., Gallagher P. M., O'Neill S. J., McElvaney N. G. (2001) J. Biol. Chem. 276, 35494–35499 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
