

Control of lipopolysaccharide biosynthesis by FtsH-mediated proteolysis of LpxC is conserved in enterobacteria but not in all gram-negative bacteria
- PMID: 21193611
- PMCID: PMC3067583
- DOI: 10.1128/JB.01043-10
Control of lipopolysaccharide biosynthesis by FtsH-mediated proteolysis of LpxC is conserved in enterobacteria but not in all gram-negative bacteria
Abstract
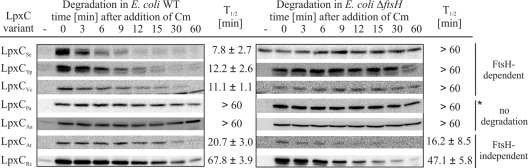
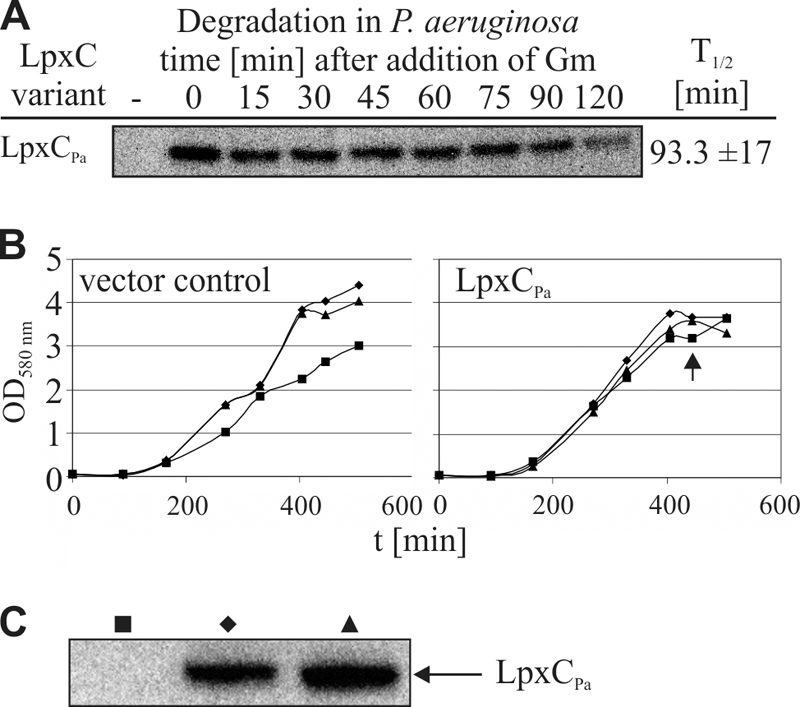
Despite the essential function of lipopolysaccharides (LPS) in Gram-negative bacteria, it is largely unknown how the exact amount of this molecule in the outer membrane is controlled. The first committed step in LPS biosynthesis is catalyzed by the LpxC enzyme. In Escherichia coli, the cellular concentration of LpxC is adjusted by the only essential protease in this organism, the membrane-anchored metalloprotease FtsH. Turnover of E. coli LpxC requires a length- and sequence-specific C-terminal degradation signal. LpxC proteins from Salmonella, Yersinia, and Vibrio species carry similar C-terminal ends and, like the E. coli enzyme, were degraded by FtsH. Although LpxC proteins are highly conserved in Gram-negative bacteria, there are striking differences in their C termini. The Aquifex aeolicus enzyme, which is devoid of the C-terminal extension, was stable in E. coli, whereas LpxC from the alphaproteobacteria Agrobacterium tumefaciens and Rhodobacter capsulatus was degraded by the Lon protease. Proteolysis of the A. tumefaciens protein required the C-terminal end of LpxC. High stability of Pseudomonas aeruginosa LpxC in E. coli and P. aeruginosa suggested that Pseudomonas uses a proteolysis-independent strategy to control its LPS content. The differences in LpxC turnover along with previously reported differences in susceptibility against antimicrobial compounds have important implications for the potential of LpxC as a drug target.
Figures





Similar articles
-
An Essential Membrane Protein Modulates the Proteolysis of LpxC to Control Lipopolysaccharide Synthesis in Escherichia coli.mBio. 2020 May 19;11(3):e00939-20. doi: 10.1128/mBio.00939-20. mBio. 2020. PMID: 32430473 Free PMC article.
-
FtsH-mediated coordination of lipopolysaccharide biosynthesis in Escherichia coli correlates with the growth rate and the alarmone (p)ppGpp.J Bacteriol. 2013 May;195(9):1912-9. doi: 10.1128/JB.02134-12. Epub 2013 Feb 15. J Bacteriol. 2013. PMID: 23417489 Free PMC article.
-
The C-terminal end of LpxC is required for degradation by the FtsH protease.Mol Microbiol. 2006 Feb;59(3):1025-36. doi: 10.1111/j.1365-2958.2005.04994.x. Mol Microbiol. 2006. PMID: 16420369
-
Antibacterial Drug Discovery Targeting the Lipopolysaccharide Biosynthetic Enzyme LpxC.Cold Spring Harb Perspect Med. 2016 Jul 1;6(7):a025304. doi: 10.1101/cshperspect.a025304. Cold Spring Harb Perspect Med. 2016. PMID: 27235477 Free PMC article. Review.
-
Checkpoints That Regulate Balanced Biosynthesis of Lipopolysaccharide and Its Essentiality in Escherichia coli.Int J Mol Sci. 2021 Dec 24;23(1):189. doi: 10.3390/ijms23010189. Int J Mol Sci. 2021. PMID: 35008618 Free PMC article. Review.
Cited by
-
Mechanisms decreasing in vitro susceptibility to the LpxC inhibitor CHIR-090 in the gram-negative pathogen Pseudomonas aeruginosa.Antimicrob Agents Chemother. 2012 Jan;56(1):17-27. doi: 10.1128/AAC.05417-11. Epub 2011 Oct 24. Antimicrob Agents Chemother. 2012. PMID: 22024823 Free PMC article.
-
The Escherichia coli Phospholipase PldA Regulates Outer Membrane Homeostasis via Lipid Signaling.mBio. 2018 Mar 20;9(2):e00379-18. doi: 10.1128/mBio.00379-18. mBio. 2018. PMID: 29559571 Free PMC article.
-
Heat-shock proteases promote survival of Pseudomonas aeruginosa during growth arrest.Proc Natl Acad Sci U S A. 2020 Feb 25;117(8):4358-4367. doi: 10.1073/pnas.1912082117. Epub 2020 Feb 6. Proc Natl Acad Sci U S A. 2020. PMID: 32029587 Free PMC article.
-
LpxC inhibitors as new antibacterial agents and tools for studying regulation of lipid A biosynthesis in Gram-negative pathogens.mBio. 2014 Sep 30;5(5):e01551-14. doi: 10.1128/mBio.01551-14. mBio. 2014. PMID: 25271285 Free PMC article.
-
Signaling through the Salmonella PbgA-LapB regulatory complex activates LpxC proteolysis and limits lipopolysaccharide biogenesis during stationary-phase growth.J Bacteriol. 2024 Apr 18;206(4):e0030823. doi: 10.1128/jb.00308-23. Epub 2024 Mar 27. J Bacteriol. 2024. PMID: 38534107 Free PMC article.
References
-
- Barembruch, C., and R. Hengge. 2007. Cellular levels and activity of the flagellar sigma factor FliA of Escherichia coli are controlled by FlgM-modulated proteolysis. Mol. Microbiol. 65:76-89. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
