

Bacterial effector HopF2 suppresses arabidopsis innate immunity at the plasma membrane
- PMID: 21198360
- PMCID: PMC3071429
- DOI: 10.1094/MPMI-07-10-0150
Bacterial effector HopF2 suppresses arabidopsis innate immunity at the plasma membrane
Abstract
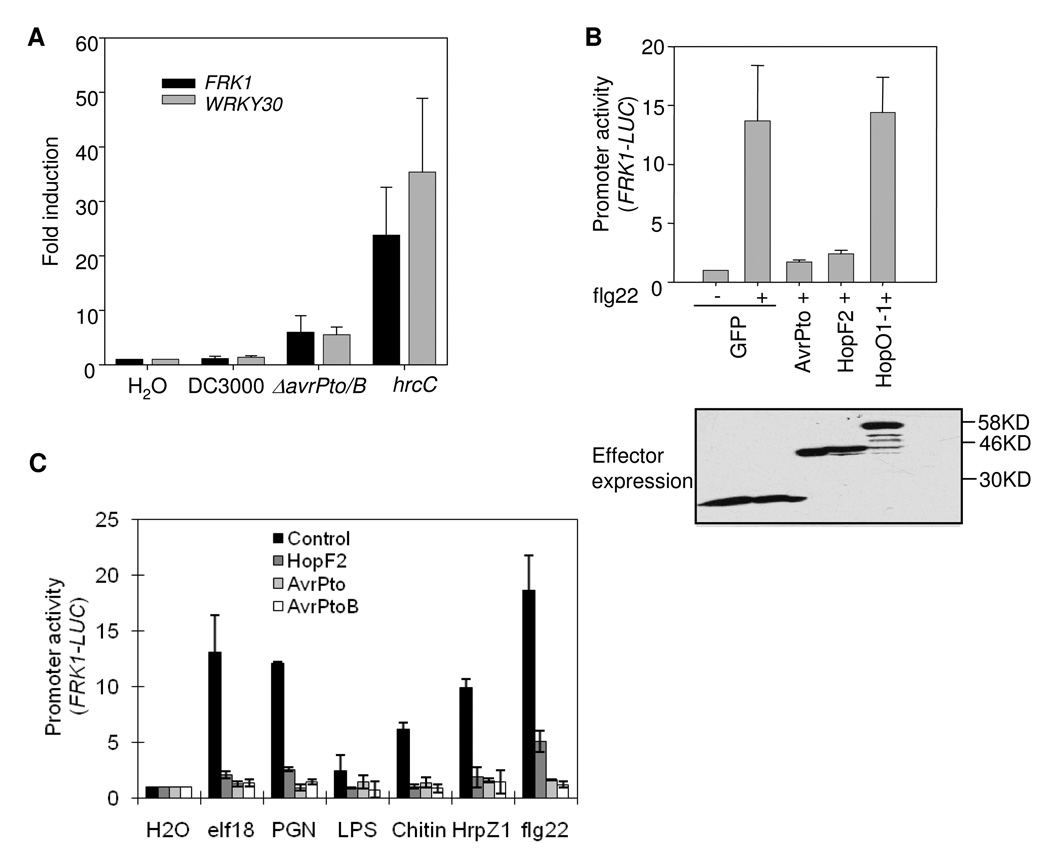
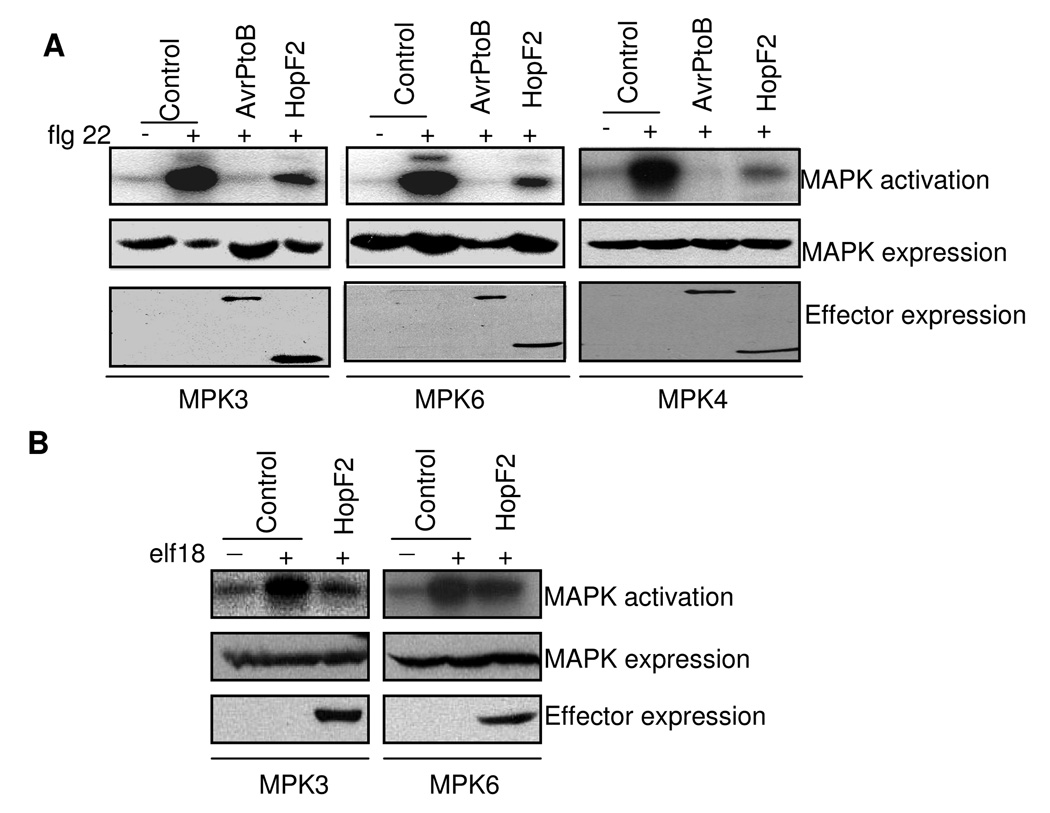
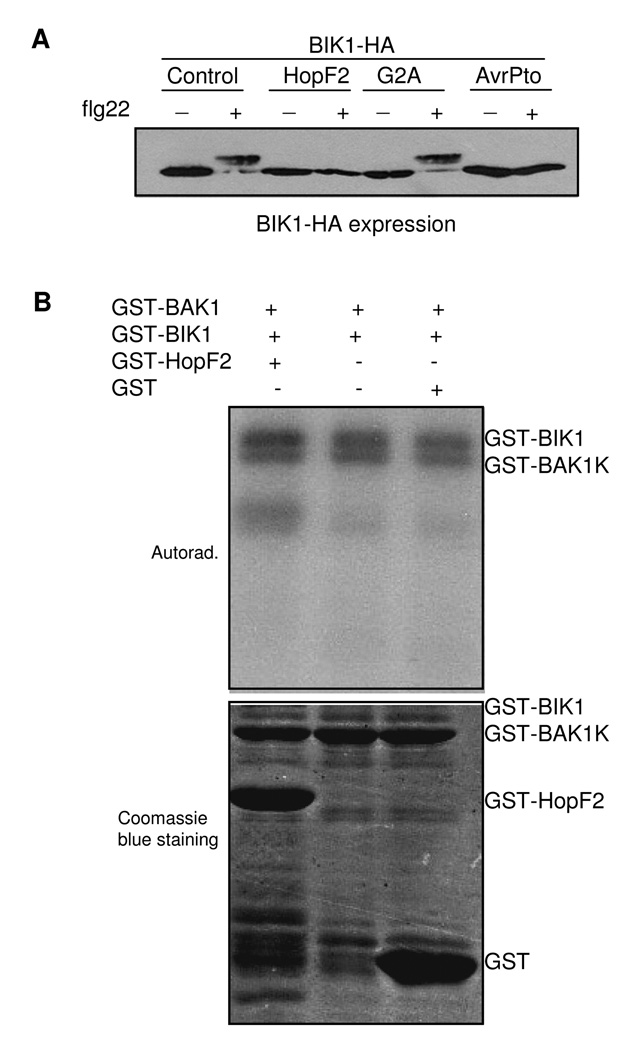
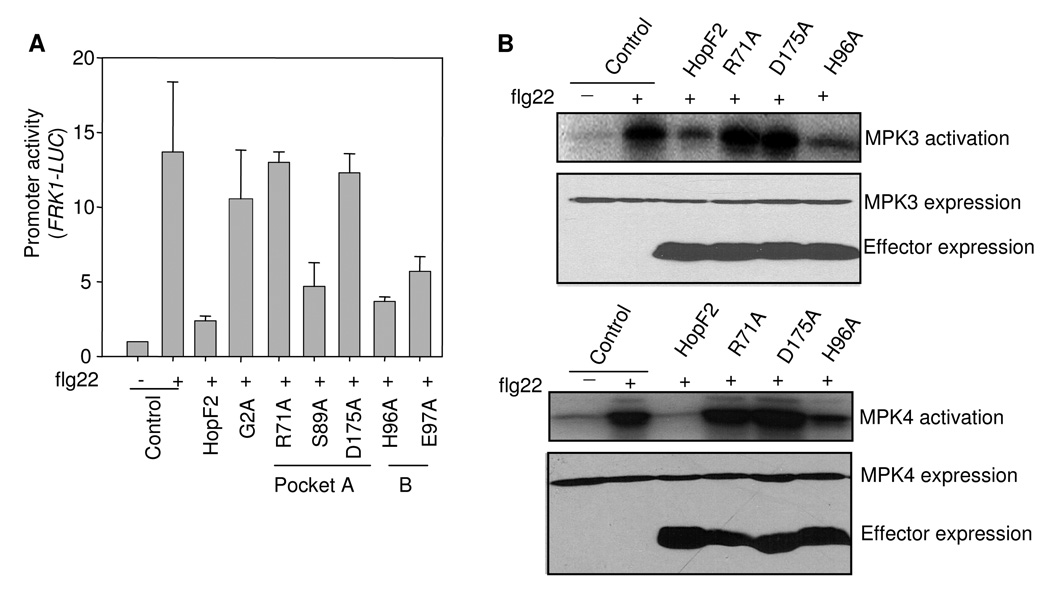
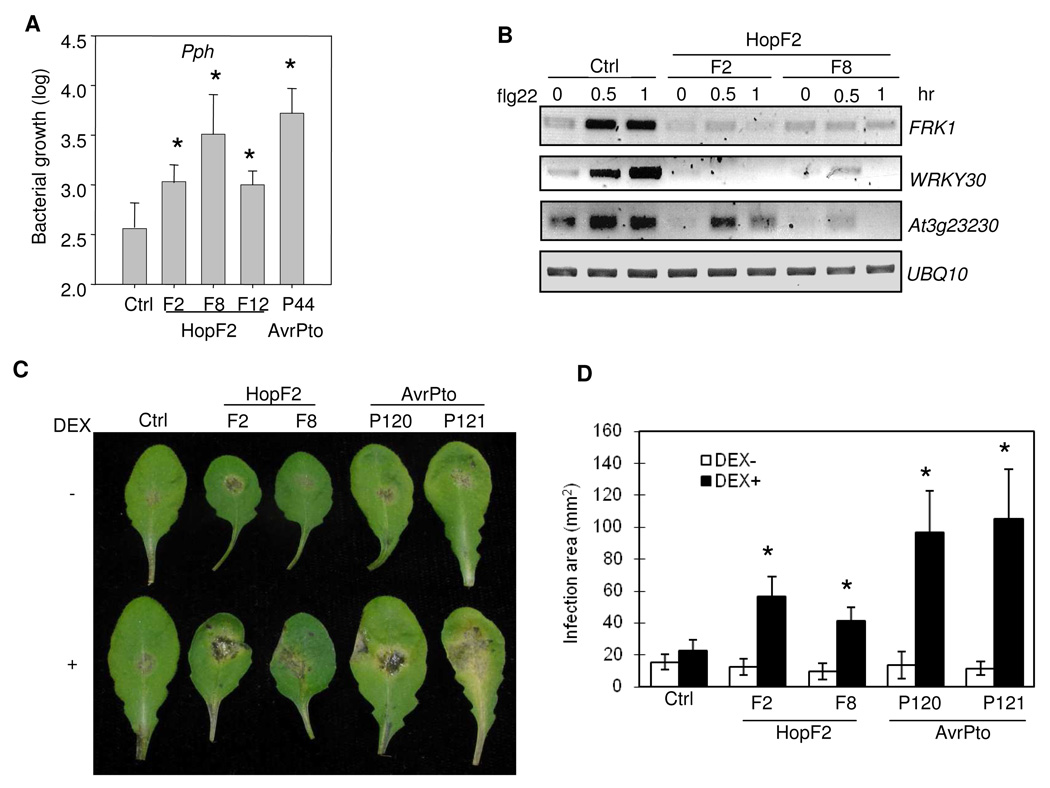
Many bacterial pathogens inject a cocktail of effector proteins into host cells through type III secretion systems. These effectors act in concert to modulate host physiology and immune signaling, thereby promoting pathogenicity. In a search for additional Pseudomonas syringae effectors in suppressing plant innate immunity triggered by pathogen or microbe-associated molecular patterns (PAMPs or MAMPs), we identified P. syringae tomato DC3000 effector HopF2 as a potent suppressor of early immune-response gene transcription and mitogen-activated protein kinase (MAPK) signaling activated by multiple MAMPs, including bacterial flagellin, elongation factor Tu, peptidoglycan, lipopolysaccharide and HrpZ1 harpin, and fungal chitin. The conserved surface-exposed residues of HopF2 are essential for its MAMP suppression activity. HopF2 is targeted to the plant plasma membrane through a putative myristoylation site, and the membrane association appears to be required for its MAMP-suppression function. Expression of HopF2 in plants potently diminished the flagellin-induced phosphorylation of BIK1, a plasma membrane-associated cytoplasmic kinase that is rapidly phosphorylated within one minute upon flagellin perception. Thus, HopF2 likely intercepts MAMP signaling at the plasma membrane immediately of signal perception. Consistent with the potent suppression function of multiple MAMP signaling, expression of HopF2 in transgenic plants compromised plant nonhost immunity to bacteria P. syringae pv. Phaseolicola and plant immunity to the necrotrophic fungal pathogen Botrytis cinerea.
Figures
Similar articles
-
The Pseudomonas syringae effector HopF2 suppresses Arabidopsis immunity by targeting BAK1.Plant J. 2014 Jan;77(2):235-45. doi: 10.1111/tpj.12381. Epub 2013 Dec 9. Plant J. 2014. PMID: 24237140 Free PMC article.
-
The majority of the type III effector inventory of Pseudomonas syringae pv. tomato DC3000 can suppress plant immunity.Mol Plant Microbe Interact. 2009 Sep;22(9):1069-80. doi: 10.1094/MPMI-22-9-1069. Mol Plant Microbe Interact. 2009. PMID: 19656042 Free PMC article.
-
Plant immunity directly or indirectly restricts the injection of type III effectors by the Pseudomonas syringae type III secretion system.Plant Physiol. 2010 Sep;154(1):233-44. doi: 10.1104/pp.110.159723. Epub 2010 Jul 12. Plant Physiol. 2010. PMID: 20624999 Free PMC article.
-
Responses of Arabidopsis thaliana to challenge by Pseudomonas syringae.Mol Cells. 2008 May 31;25(3):323-31. Epub 2008 May 16. Mol Cells. 2008. PMID: 18483469 Review.
-
Defining essential processes in plant pathogenesis with Pseudomonas syringae pv. tomato DC3000 disarmed polymutants and a subset of key type III effectors.Mol Plant Pathol. 2018 Jul;19(7):1779-1794. doi: 10.1111/mpp.12655. Epub 2018 Feb 1. Mol Plant Pathol. 2018. PMID: 29277959 Free PMC article. Review.
Cited by
-
Pseudomonas syringae: what it takes to be a pathogen.Nat Rev Microbiol. 2018 May;16(5):316-328. doi: 10.1038/nrmicro.2018.17. Epub 2018 Feb 26. Nat Rev Microbiol. 2018. PMID: 29479077 Free PMC article. Review.
-
Suppression of NLR-mediated plant immune detection by bacterial pathogens.J Exp Bot. 2023 Oct 13;74(19):6069-6088. doi: 10.1093/jxb/erad246. J Exp Bot. 2023. PMID: 37429579 Free PMC article. Review.
-
The leucine-rich repeat receptor kinase QSK1 regulates PRR-RBOHD complexes targeted by the bacterial effector HopF2Pto.Plant Cell. 2024 Oct 21;36(12):4932-51. doi: 10.1093/plcell/koae267. Online ahead of print. Plant Cell. 2024. PMID: 39431742 Free PMC article.
-
Metaeffector interactions modulate the type III effector-triggered immunity load of Pseudomonas syringae.PLoS Pathog. 2022 May 16;18(5):e1010541. doi: 10.1371/journal.ppat.1010541. eCollection 2022 May. PLoS Pathog. 2022. PMID: 35576228 Free PMC article.
-
Multiple host targets of Pseudomonas effector protein HopM1 form a protein complex regulating apoplastic immunity and water homeostasis.bioRxiv [Preprint]. 2023 Aug 1:2023.07.31.551310. doi: 10.1101/2023.07.31.551310. bioRxiv. 2023. PMID: 37577537 Free PMC article. Preprint.
References
-
- Asai T, Tena G, Plotnikova J, Willmann MR, Chiu WL, Gomez-Gomez L, Boller T, Ausubel FM, Sheen J. MAP kinase signalling cascade in Arabidopsis innate immunity. Nature. 2002;415:977–983. - PubMed
-
- Ausubel FM. Are innate immune signaling pathways in plants and animals conserved? Nat Immunol. 2005;6:973–979. - PubMed
-
- Axtell MJ, Staskawicz BJ. Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell. 2003;112:369–377. - PubMed
-
- Boller T, Felix G. A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol. 2009;60:379–406. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
