

Characterization of immortalized mesenchymal stem cells derived from foetal porcine pancreas
- PMID: 21199007
- PMCID: PMC6496693
- DOI: 10.1111/j.1365-2184.2010.00714.x
Characterization of immortalized mesenchymal stem cells derived from foetal porcine pancreas
Abstract
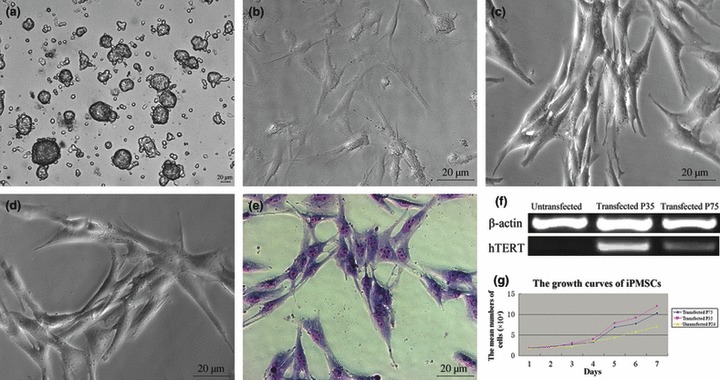
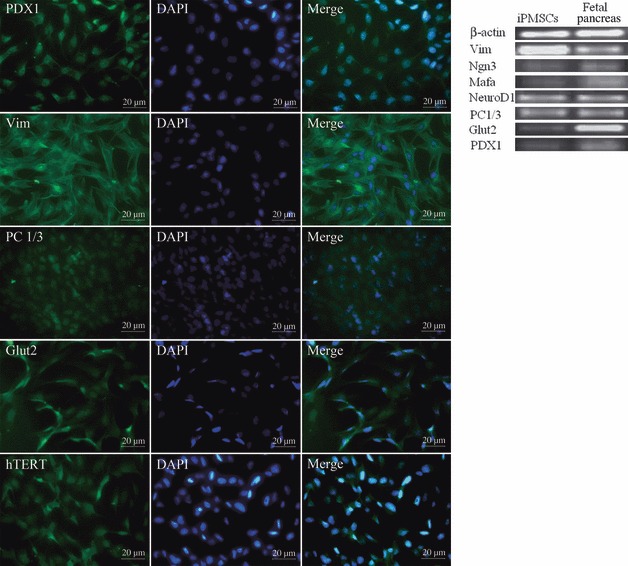
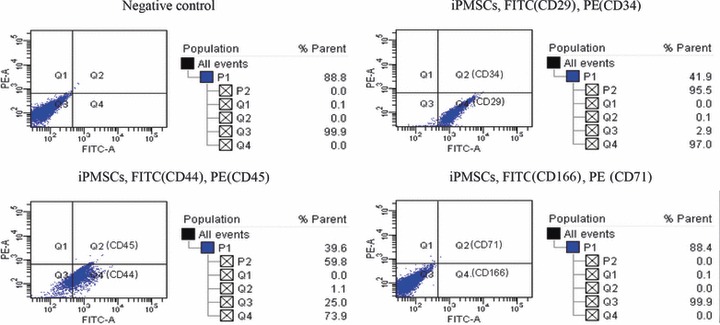
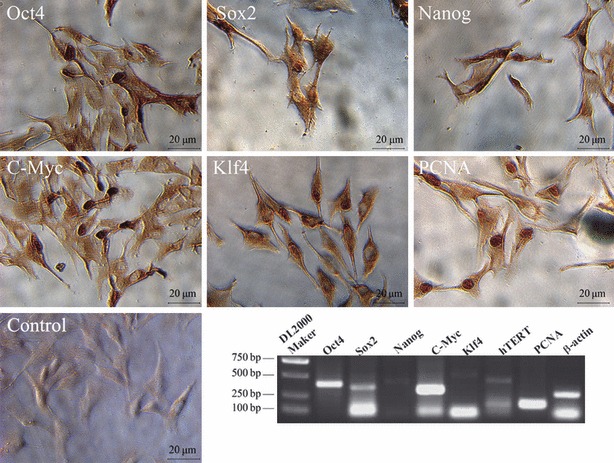
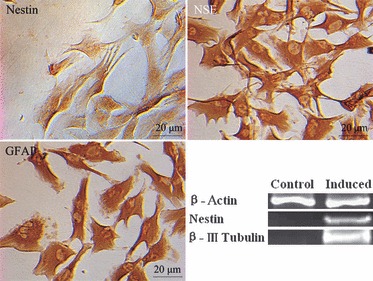
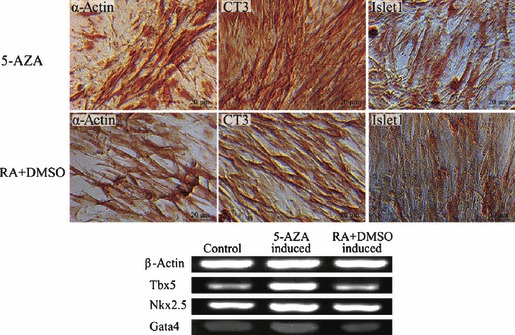
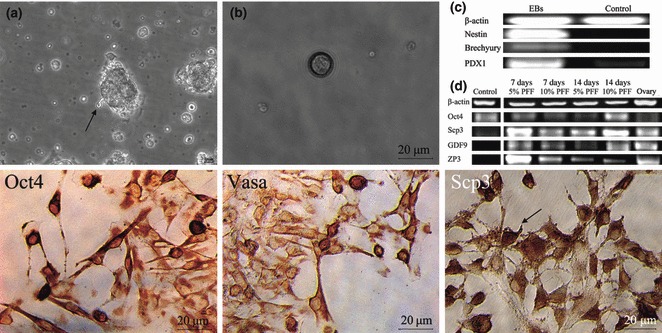
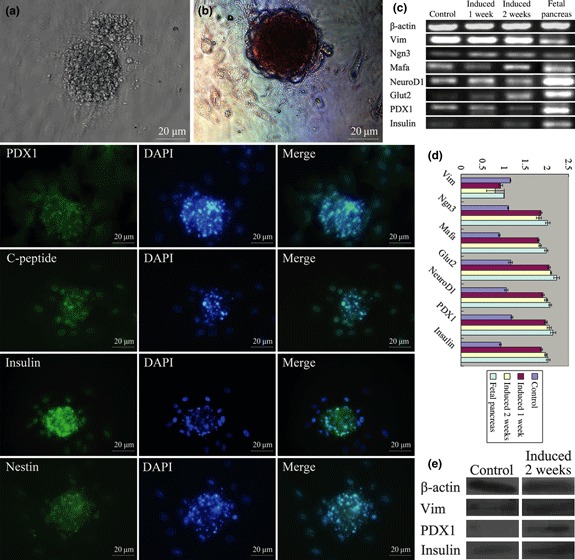
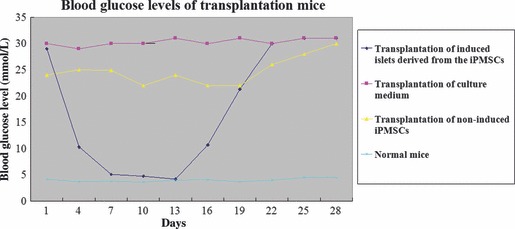
Islet replacement therapy is limited by shortage of donor islet cells. Usage of islet cells derived from porcine pancreatic stem cells (PSCs) is currently viewed as the most promising alternative for human islet transplantation. However, PSCs are rare and have a finite proliferative lifespan. In this study, we isolated and established an immortalized mesenchymal stem cell (MSC) line derived from foetal porcine pancreas, by transfecting human telomerase reverse transcriptase (hTERT) and called these immortalized pancreatic mesenchymal stem cells (iPMSCs). The iPMSCs have been cultured for more than 80 passages and have capacity to differentiate into neurons, cardiomyocytes, germ cells and islet-like cells, analysed by morphology, RT-PCR, western blotting, immunofluorescence, immunocytochemistry and transplantation assay. Islets derived from iPMSCs reversed hyperglycaemia in streptozotocin-induced diabetic mice and secreted insulin and C-peptide in vitro. These results demonstrated that iPMSCs might provide unlimited resources for islet replacement therapy and models for functional cell differentiation.
© 2010 Blackwell Publishing Ltd.
Figures
Similar articles
-
Pancreatic islet-like clusters from bone marrow mesenchymal stem cells of human first-trimester abortus can cure streptozocin-induced mouse diabetes.Rejuvenation Res. 2010 Dec;13(6):695-706. doi: 10.1089/rej.2009.1016. Epub 2011 Jan 4. Rejuvenation Res. 2010. PMID: 21204652
-
Reduction of marginal mass required for successful islet transplantation in a diabetic rat model using adipose tissue-derived mesenchymal stromal cells.Cytotherapy. 2018 Sep;20(9):1124-1142. doi: 10.1016/j.jcyt.2018.06.001. Epub 2018 Jul 29. Cytotherapy. 2018. PMID: 30068495
-
Establishing a human pancreatic stem cell line and transplanting induced pancreatic islets to reverse experimental diabetes in rats.Sci China C Life Sci. 2008 Sep;51(9):779-88. doi: 10.1007/s11427-008-0109-6. Epub 2008 Aug 24. Sci China C Life Sci. 2008. PMID: 18726523
-
Generation of pancreatic islet cells from human embryonic stem cells.Sci China C Life Sci. 2009 Jul;52(7):615-21. doi: 10.1007/s11427-009-0095-3. Epub 2009 Jul 30. Sci China C Life Sci. 2009. PMID: 19641866 Review.
-
Animal models to study adult stem cell-derived, in vitro-generated islet implantation.ILAR J. 2004;45(3):259-67. doi: 10.1093/ilar.45.3.259. ILAR J. 2004. PMID: 15229373 Review.
Cited by
-
Meta-analysis of the Mesenchymal Stem Cells Immortalization Protocols: A Guideline for Regenerative Medicine.Curr Stem Cell Res Ther. 2024;19(7):1009-1020. doi: 10.2174/011574888X268464231016070900. Curr Stem Cell Res Ther. 2024. PMID: 38221663
-
miR-375 controls porcine pancreatic stem cell fate by targeting 3-phosphoinositide-dependent protein kinase-1 (Pdk1).Cell Prolif. 2016 Jun;49(3):395-406. doi: 10.1111/cpr.12263. Cell Prolif. 2016. PMID: 27218665 Free PMC article.
-
Expression profile of Nanos2 gene in dairy goat and its inhibitory effect on Stra8 during meiosis.Cell Prolif. 2014 Oct;47(5):396-405. doi: 10.1111/cpr.12128. Cell Prolif. 2014. PMID: 25195564 Free PMC article.
-
Conservation and function of Dazl in promoting the meiosis of goat male germline stem cells.Mol Biol Rep. 2014 May;41(5):2697-707. doi: 10.1007/s11033-014-3156-z. Epub 2014 Jan 30. Mol Biol Rep. 2014. PMID: 24477583
-
[Construction of human mucosa oral epithelial cell lines overexpressing telomerase reverse transcriptase gene mediated by lentivirus].Hua Xi Kou Qiang Yi Xue Za Zhi. 2016 Oct 1;34(5):443-447. doi: 10.7518/hxkq.2016.05.002. Hua Xi Kou Qiang Yi Xue Za Zhi. 2016. PMID: 28326698 Free PMC article. Chinese.
References
-
- Cai J, Yu C, Liu Y, Chen S, Guo Y, Yong Jun et al. (2010) Generation of homogeneous PDX1+ pancreatic progenitors from human es cell‐derived endoderm cells. J. Mol. Cell Biol. 2, 50–60. - PubMed
-
- Steve KW, Andre BH (2006) Human Embryonic stem cells: technological challenges towards therapy. Clin. Exp. Pharmacol. Physiol. 33, 489–495. - PubMed
-
- Rogers SA, Chen F, Talcott MR, Faulkner C, Thomas JM, Thevis M et al. (2007) Long‐term engraftment following transplantation of pig pancreatic primordia into non‐immunosuppressed diabetic rhesus macaques. Xenotransplantation 14, 591–560. - PubMed
-
- Feng R, Zhang H, Wang Y, Qiao H, Zhao T, Shen W et al. (2007) Isolation, culture and induced differentiation of fetal porcine islet derived pancreatic stem cell. Agric. Sci. China 6, 742–748.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
