

Subversion of innate defenses by the interplay between DENV and pre-existing enhancing antibodies: TLRs signaling collapse
- PMID: 21200427
- PMCID: PMC3006139
- DOI: 10.1371/journal.pntd.0000924
Subversion of innate defenses by the interplay between DENV and pre-existing enhancing antibodies: TLRs signaling collapse
Abstract
Background: The phenomenon of antibody dependent enhancement as a major determinant that exacerbates disease severity in DENV infections is well accepted. While the detailed mechanism of antibody enhanced disease severity is unclear, evidence suggests that it is associated with both increased DENV infectivity and suppression of the type I IFN and pro-inflammatory cytokine responses. Therefore, it is imperative for us to understand the intracellular mechanisms altered during ADE infection to decipher the mechanism of severe pathogenesis.
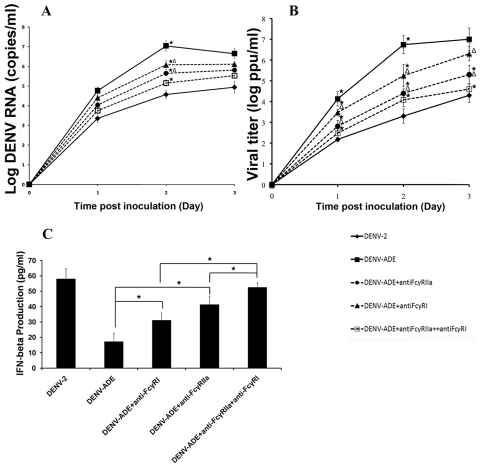
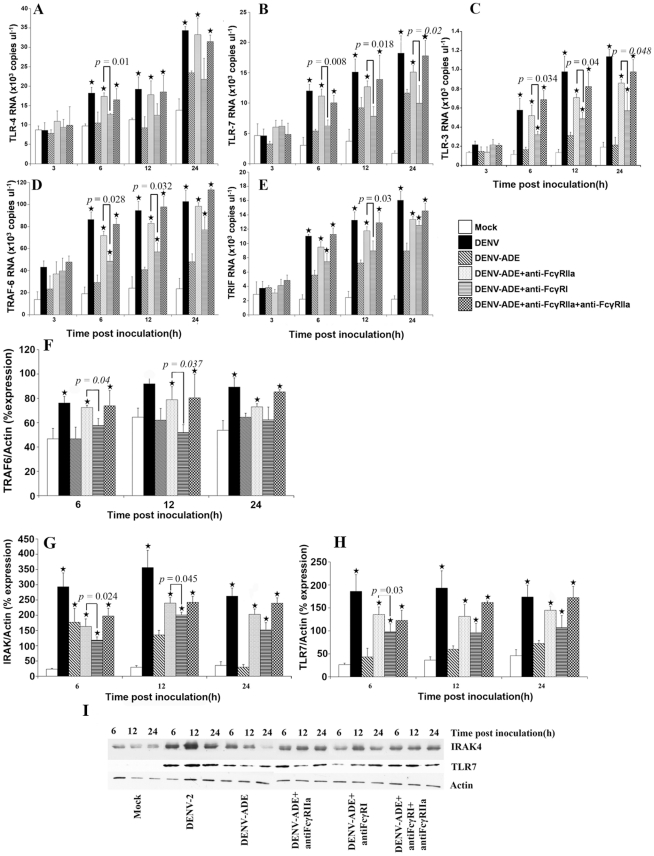
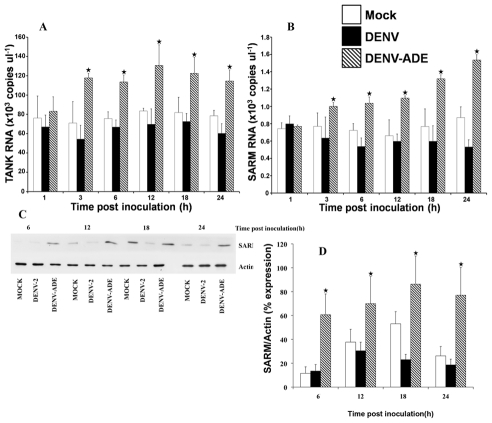
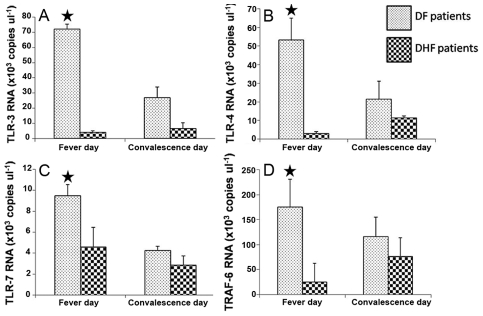
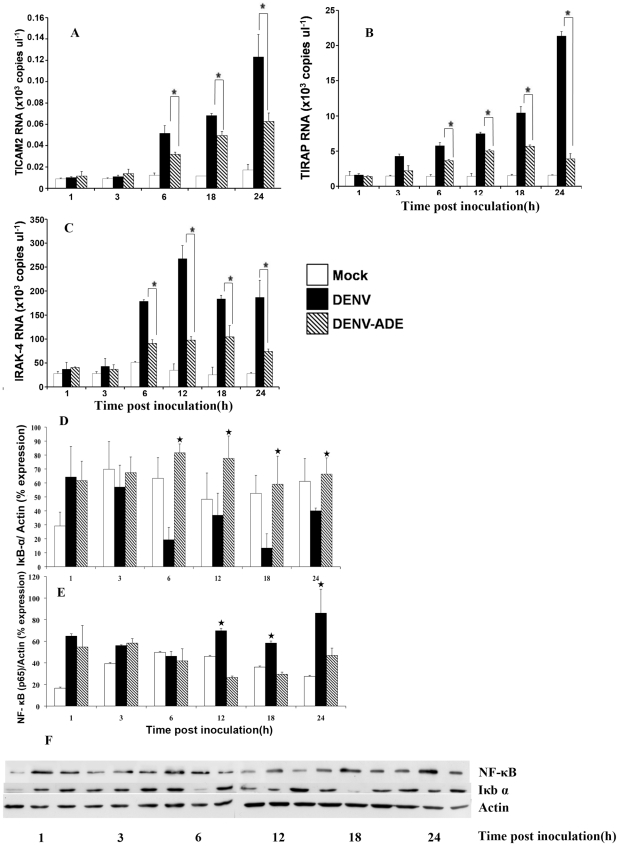
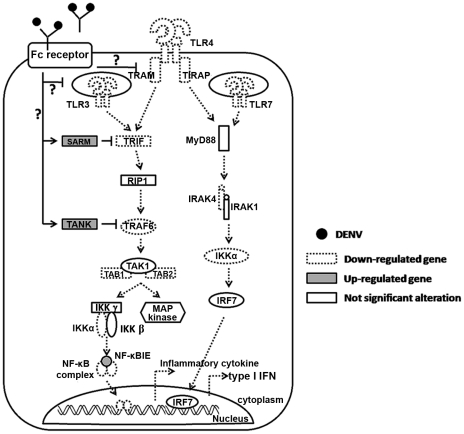
Methodology/principal findings: In this present work, qRT-PCR, immunoblotting and gene array analysis were conducted to determine whether DENV-antibody complex infection exerts a suppressive effect on the expression and/or function of the pathogen recognition patterns, focusing on the TLR-signaling pathway. We show here that FcγRI and FcγRIIa synergistically facilitated entry of DENV-antibody complexes into monocytic THP-1 cells. Ligation between DENV-antibody complexes and FcR not only down regulated TLRs gene expression but also up regulated SARM, TANK, and negative regulators of the NF-κB pathway, resulting in suppression of innate responses but increased viral production. These results were confirmed by blocking with anti-FcγRI or anti-FcγRIIa antibodies which reduced viral production, up-regulated IFN-β synthesis, and increased gene expression in the TLR-dependent signaling pathway. The negative impact of DENV-ADE infection on the TLR-dependent pathway was strongly supported by gene array screening which revealed that both MyD88-dependent and -independent signaling molecules were down regulated during DENV-ADE infection. Importantly, the same phenomenon was seen in PBMC of secondary DHF/DSS patients but not in PBMC of DF patients.
Conclusions/significance: Our present work demonstrates the mechanism by which DENV uses pre-existing immune mediators to defeat the principal activating pathway of innate defense resulting in suppression of an array of innate immune responses. Interestingly, this phenomenon specifically occurred during the severe form of DENV infection but not in the mild form of disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Gubler DJ. Epidemic dengue/dengue hemorrhagic fever as a public health, social and economic problem in the 21st century. Trends Microbiol. 2002;10:100–103. - PubMed
-
- Eradication WHOCDCPa. 2000. Strengthening implementation of the global strategy for dengue fever/dengue haemorrhagic fever prevention and control : report of the informal consultation. 21.
-
- Chaturvedi UC, Agarwal R, Elbishbishi EA, Mustafa AS. Cytokine cascade in dengue hemorrhagic fever: implications for pathogenesis. FEMS Immunol Med Microbiol. 2000;28:183–188. - PubMed
-
- Chaturvedi UC, Shrivastava R, Tripathi RK, Nagar R. Dengue virus-specific suppressor T cells: current perspectives. FEMS Immunol Med Microbiol. 2007;50:285–299. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
