

Critical role of macrophages and their activation via MyD88-NFκB signaling in lung innate immunity to Mycoplasma pneumoniae
- PMID: 21203444
- PMCID: PMC3009709
- DOI: 10.1371/journal.pone.0014417
Critical role of macrophages and their activation via MyD88-NFκB signaling in lung innate immunity to Mycoplasma pneumoniae
Abstract
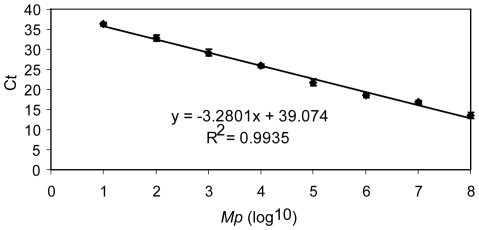
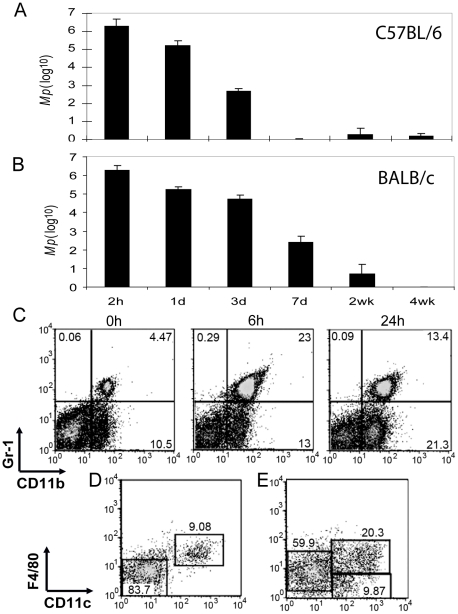
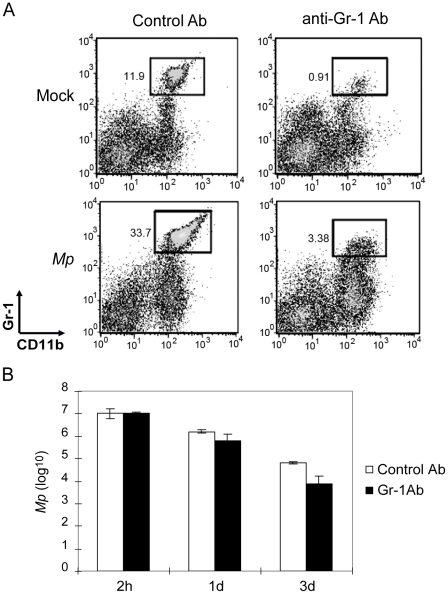
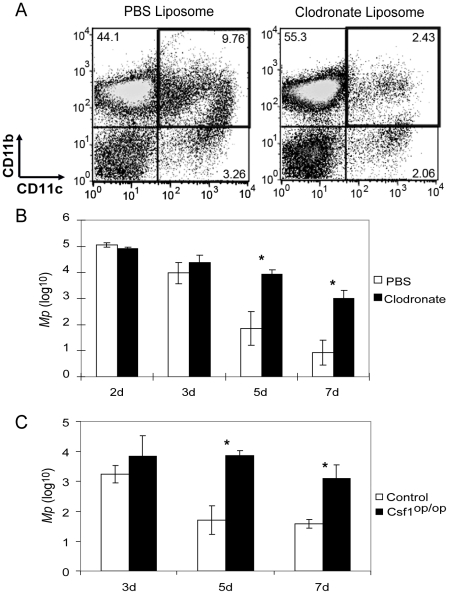
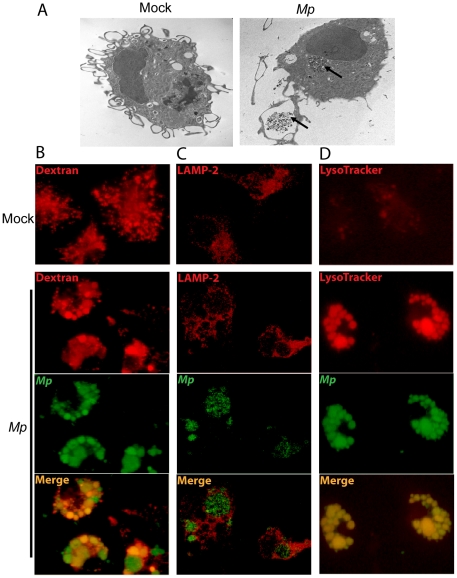
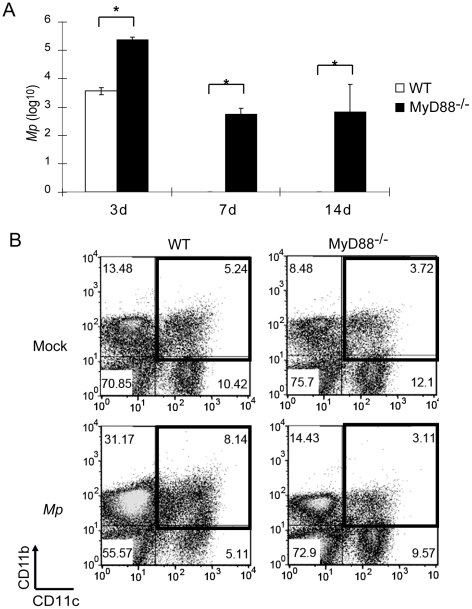
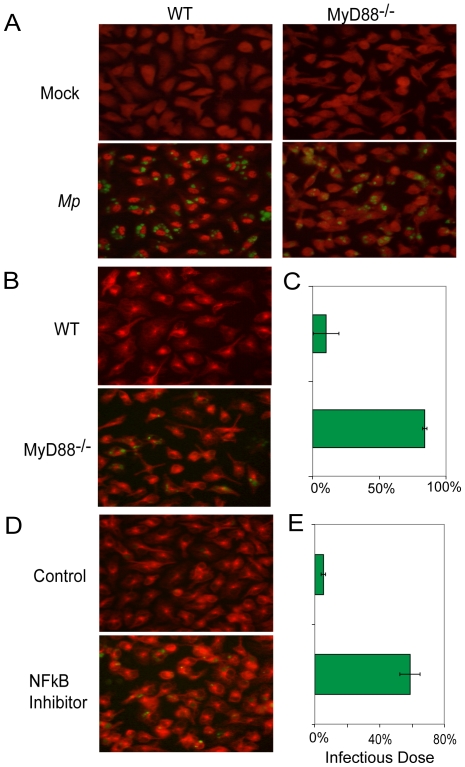
Mycoplasma pneumoniae (Mp), a common cause of pneumonia, is associated with asthma; however, the mechanisms underlying this association remain unclear. We investigated the cellular immune response to Mp in mice. Intranasal inoculation with Mp elicited infiltration of the lungs with neutrophils, monocytes and macrophages. Systemic depletion of macrophages, but not neutrophils, resulted in impaired clearance of Mp from the lungs. Accumulation and activation of macrophages were decreased in the lungs of MyD88(-/-) mice and clearance of Mp was impaired, indicating that MyD88 is a key signaling protein in the anti-Mp response. MyD88-dependent signaling was also required for the Mp-induced activation of NFκB, which was essential for macrophages to eliminate the microbe in vitro. Thus, MyD88-NFκB signaling in macrophages is essential for clearance of Mp from the lungs.
Conflict of interest statement
Figures
References
-
- Sleiman PM, Hakonarson H. Recent advances in the genetics and genomics of asthma and related traits. Curr Opin Pediatr. 2010;22:307–312. - PubMed
-
- Vercelli D. Discovering susceptibility genes for asthma and allergy. Nat Rev Immunol. 2008;8:169–182. - PubMed
-
- Martin RJ. Infections and asthma. Clin Chest Med. 2006;27:87–98. - PubMed
-
- Seggev JS, Lis I, Siman-Tov R, Gutman R, Abu-Samara H, et al. Mycoplasma pneumoniae is a frequent cause of exacerbation of bronchial asthma in adults. Ann Allergy. 1986;57:263–265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
