

Circadian disruption accelerates tumor growth and angio/stromagenesis through a Wnt signaling pathway
- PMID: 21203463
- PMCID: PMC3009728
- DOI: 10.1371/journal.pone.0015330
Circadian disruption accelerates tumor growth and angio/stromagenesis through a Wnt signaling pathway
Abstract
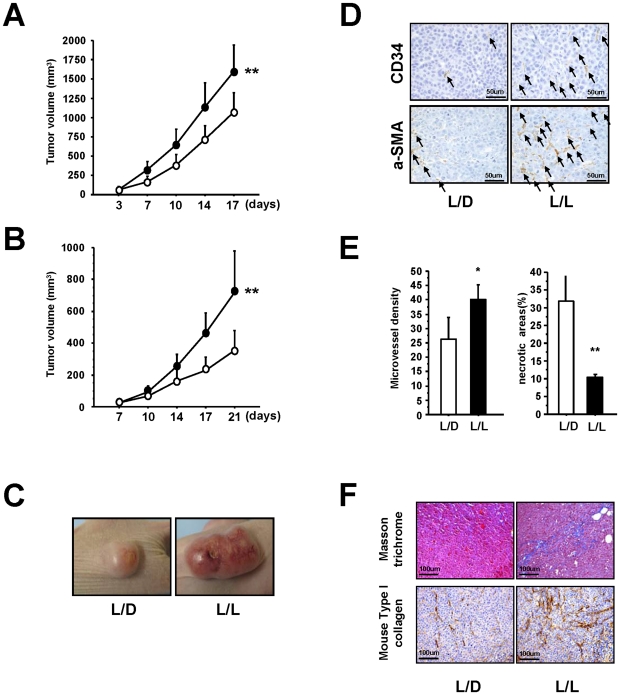
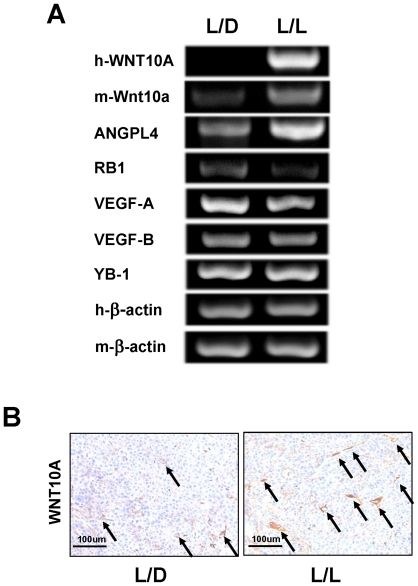
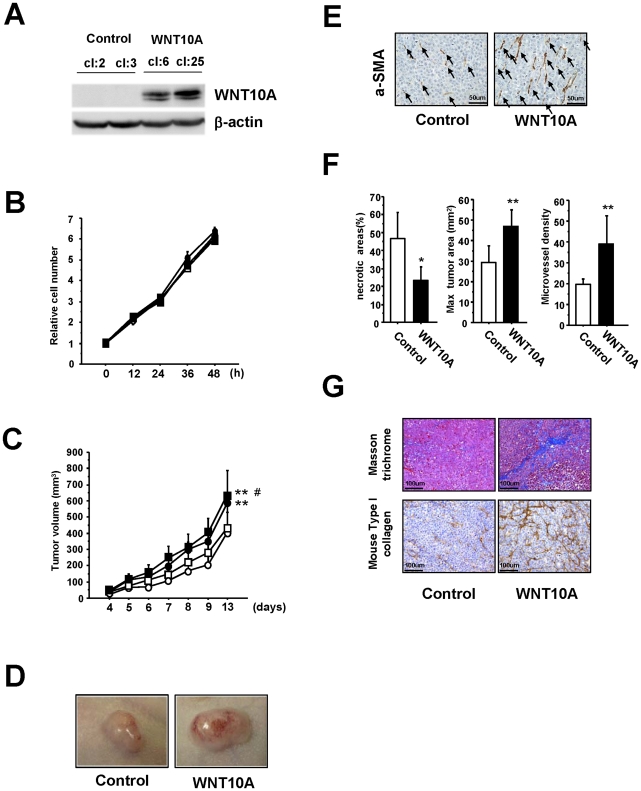
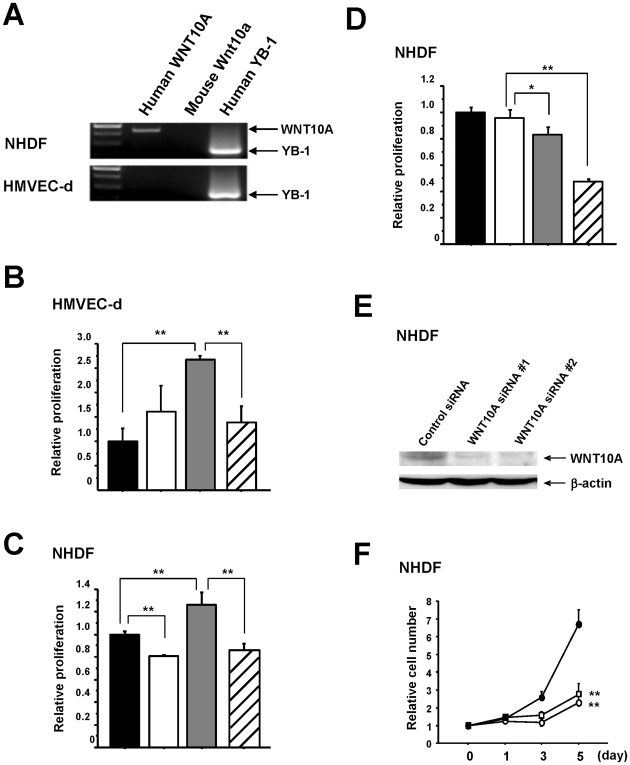
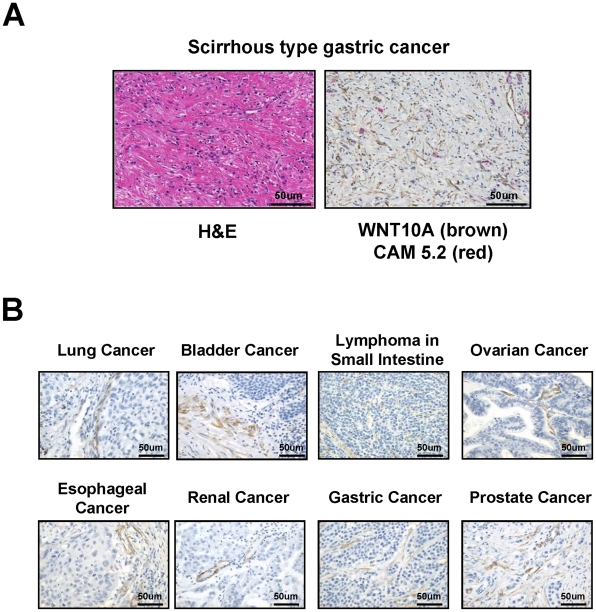
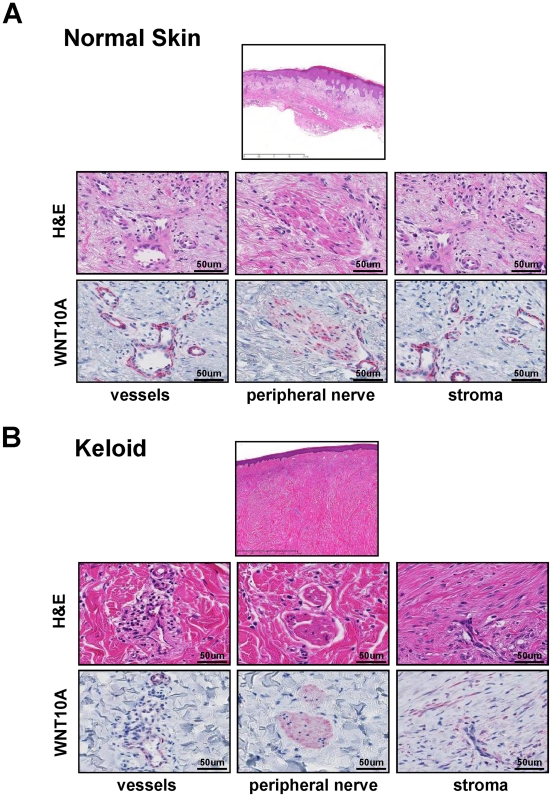
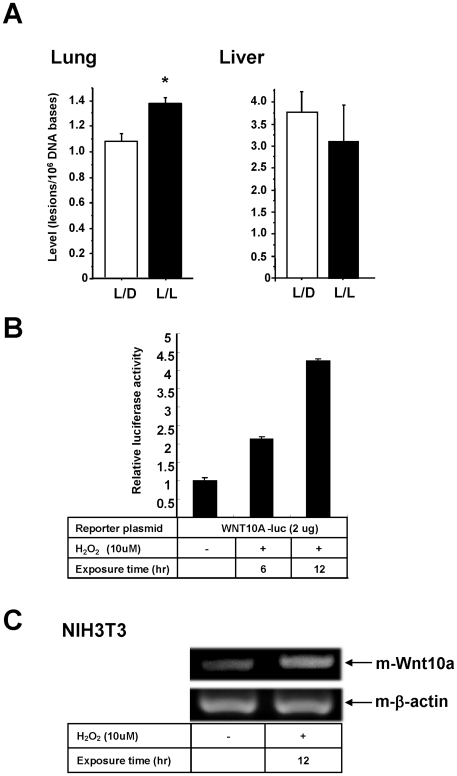
Epidemiologic studies show a high incidence of cancer in shift workers, suggesting a possible relationship between circadian rhythms and tumorigenesis. However, the precise molecular mechanism played by circadian rhythms in tumor progression is not known. To identify the possible mechanisms underlying tumor progression related to circadian rhythms, we set up nude mouse xenograft models. HeLa cells were injected in nude mice and nude mice were moved to two different cases, one case is exposed to a 24-hour light cycle (L/L), the other is a more "normal" 12-hour light/dark cycle (L/D). We found a significant increase in tumor volume in the L/L group compared with the L/D group. In addition, tumor microvessels and stroma were strongly increased in L/L mice. Although there was a hypervascularization in L/L tumors, there was no associated increase in the production of vascular endothelial cell growth factor (VEGF). DNA microarray analysis showed enhanced expression of WNT10A, and our subsequent study revealed that WNT10A stimulates the growth of both microvascular endothelial cells and fibroblasts in tumors from light-stressed mice, along with marked increases in angio/stromagenesis. Only the tumor stroma stained positive for WNT10A and WNT10A is also highly expressed in keloid dermal fibroblasts but not in normal dermal fibroblasts indicated that WNT10A may be a novel angio/stromagenic growth factor. These findings suggest that circadian disruption induces the progression of malignant tumors via a Wnt signaling pathway.
Conflict of interest statement
Figures
References
-
- Rajaratnam SM, Arendt J. Health in a 24-h society. Lancet. 2006;358:999–1005. - PubMed
-
- Navara KJ, Nelson RJ. The dark side of light at night: physiological, epidemiological, and ecological consequences. J Pineal Res. 2007;43:215–24. - PubMed
-
- Fu L, Lee CC. The circadian clock: pacemaker and tumour suppressor. Nat Rev Cancer. 2003;3:350–61. - PubMed
-
- Hansen J. Risk of breast cancer after night- and shift work: current evidence and ongoing studies in Denmark. Cancer Causes Control. 2006;17:531–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
