

Role for sumoylation in systemic inflammation and immune homeostasis in Drosophila larvae
- PMID: 21203476
- PMCID: PMC3009591
- DOI: 10.1371/journal.ppat.1001234
Role for sumoylation in systemic inflammation and immune homeostasis in Drosophila larvae
Abstract
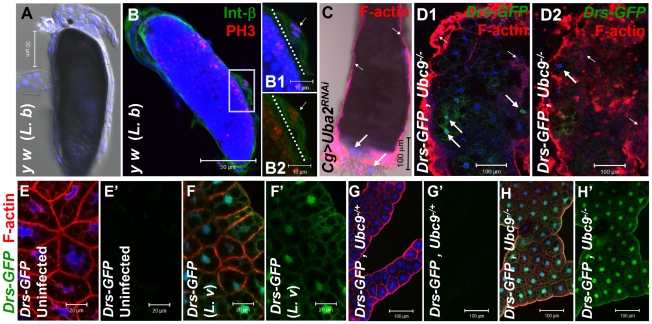
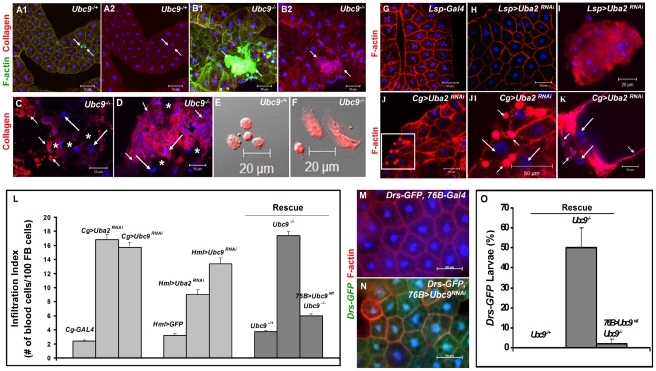
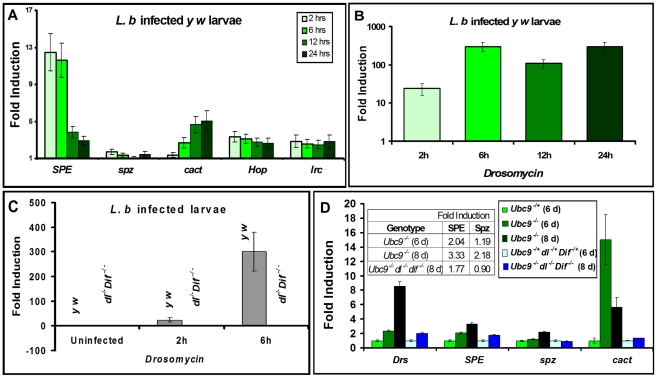
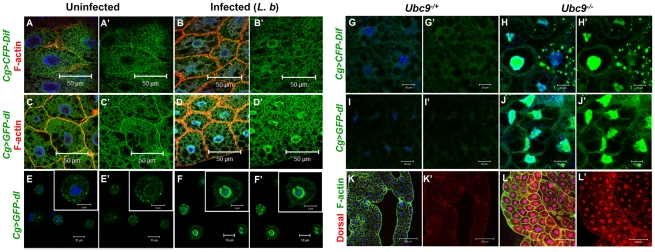
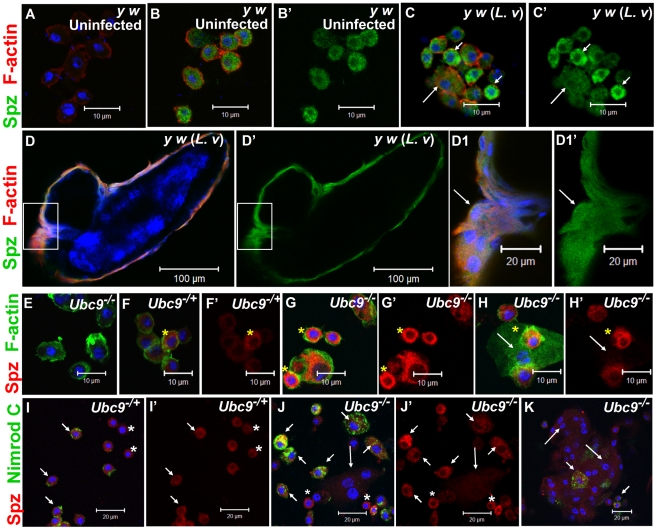
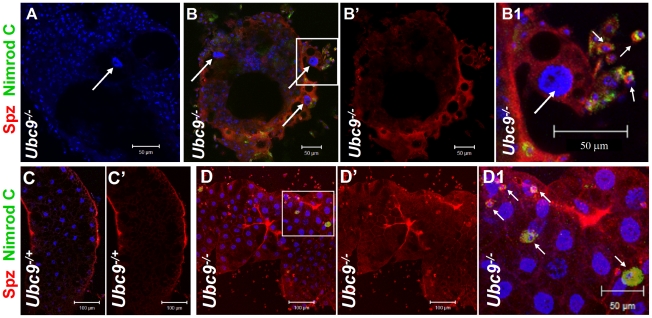
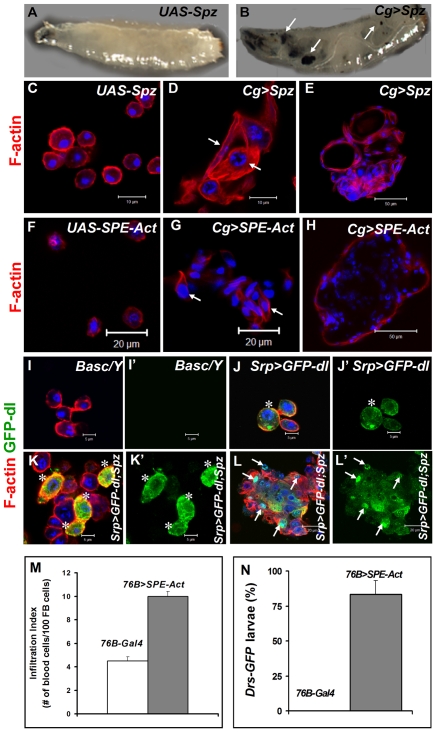
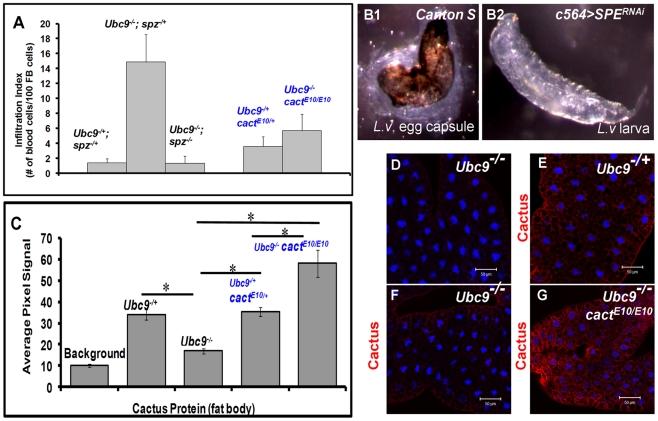
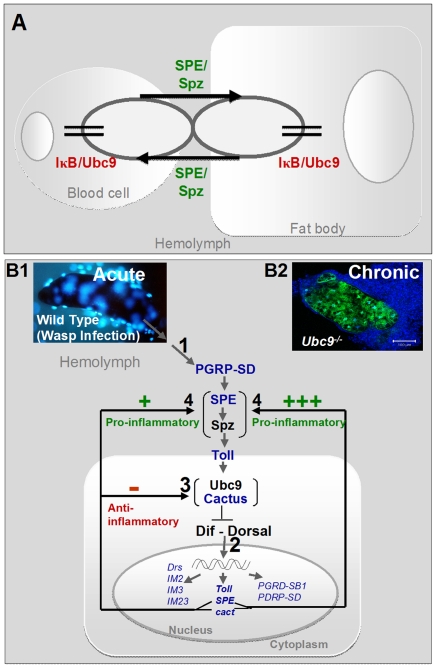
To counter systemic risk of infection by parasitic wasps, Drosophila larvae activate humoral immunity in the fat body and mount a robust cellular response resulting in encapsulation of the wasp egg. Innate immune reactions are tightly regulated and are resolved within hours. To understand the mechanisms underlying activation and resolution of the egg encapsulation response and examine if failure of the latter develops into systemic inflammatory disease, we correlated parasitic wasp-induced changes in the Drosophila larva with systemic chronic conditions in sumoylation-deficient mutants. We have previously reported that loss of either Cactus, the Drosophila (IκB) protein or Ubc9, the SUMO-conjugating enzyme, leads to constitutive activation of the humoral and cellular pathways, hematopoietic overproliferation and tumorogenesis. Here we report that parasite infection simultaneously activates NF-κB-dependent transcription of Spätzle processing enzyme (SPE) and cactus. Endogenous Spätzle protein (the Toll ligand) is expressed in immune cells and excessive SPE or Spätzle is pro-inflammatory. Consistent with this function, loss of Spz suppresses Ubc9⁻ defects. In contrast to the pro-inflammatory roles of SPE and Spätzle, Cactus and Ubc9 exert an anti-inflammatory effect. We show that Ubc9 maintains steady state levels of Cactus protein. In a series of immuno-genetic experiments, we demonstrate the existence of a robust bidirectional interaction between blood cells and the fat body and propose that wasp infection activates Toll signaling in both compartments via extracellular activation of Spätzle. Within each organ, the IκB/Ubc9-dependent inhibitory feedback resolves immune signaling and restores homeostasis. The loss of this feedback leads to chronic inflammation. Our studies not only provide an integrated framework for understanding the molecular basis of the evolutionary arms race between insect hosts and their parasites, but also offer insights into developing novel strategies for medical and agricultural pest control.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
