

A young Drosophila duplicate gene plays essential roles in spermatogenesis by regulating several Y-linked male fertility genes
- PMID: 21203494
- PMCID: PMC3009665
- DOI: 10.1371/journal.pgen.1001255
A young Drosophila duplicate gene plays essential roles in spermatogenesis by regulating several Y-linked male fertility genes
Abstract
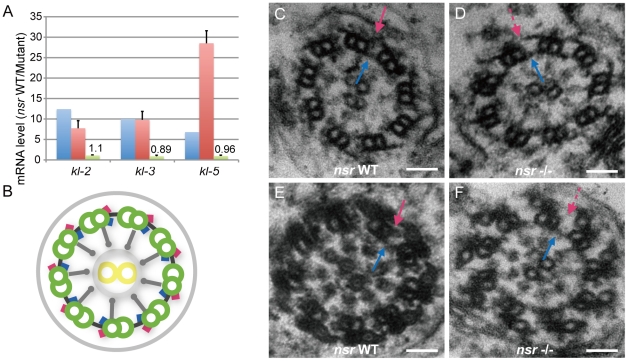
Gene duplication is supposed to be the major source for genetic innovations. However, how a new duplicate gene acquires functions by integrating into a pathway and results in adaptively important phenotypes has remained largely unknown. Here, we investigated the biological roles and the underlying molecular mechanism of the young kep1 gene family in the Drosophila melanogaster species subgroup to understand the origin and evolution of new genes with new functions. Sequence and expression analysis demonstrates that one of the new duplicates, nsr (novel spermatogenesis regulator), exhibits positive selection signals and novel subcellular localization pattern. Targeted mutagenesis and whole-transcriptome sequencing analysis provide evidence that nsr is required for male reproduction associated with sperm individualization, coiling, and structural integrity of the sperm axoneme via regulation of several Y chromosome fertility genes post-transcriptionally. The absence of nsr-like expression pattern and the presence of the corresponding cis-regulatory elements of the parental gene kep1 in the pre-duplication species Drosophila yakuba indicate that kep1 might not be ancestrally required for male functions and that nsr possibly has experienced the neofunctionalization process, facilitated by changes of trans-regulatory repertories. These findings not only present a comprehensive picture about the evolution of a new duplicate gene but also show that recently originated duplicate genes can acquire multiple biological roles and establish novel functional pathways by regulating essential genes.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Ohno S. New York: Springer-Verlag; 1970. Evolution by gene duplication.
-
- Zhou Q, Wang W. On the origin and evolution of new genes–a genomic and experimental perspective. J Genet Genomics. 2008;35:639–648. - PubMed
-
- Long M, Betran E, Thornton K, Wang W. The origin of new genes: glimpses from the young and old. Nat Rev Genet. 2003;4:865–875. - PubMed
-
- Lynch M, Conery JS. The evolutionary fate and consequences of duplicate genes. Science. 2000;290:1151–1155. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
