

Praf2 is a novel Bcl-xL/Bcl-2 interacting protein with the ability to modulate survival of cancer cells
- PMID: 21203533
- PMCID: PMC3006391
- DOI: 10.1371/journal.pone.0015636
Praf2 is a novel Bcl-xL/Bcl-2 interacting protein with the ability to modulate survival of cancer cells
Abstract
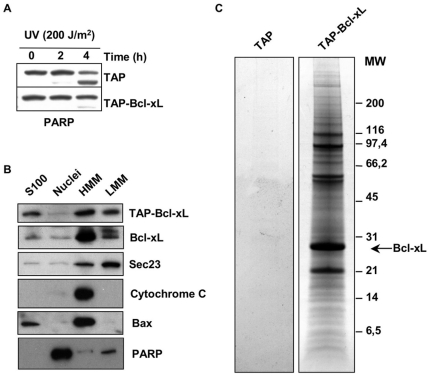
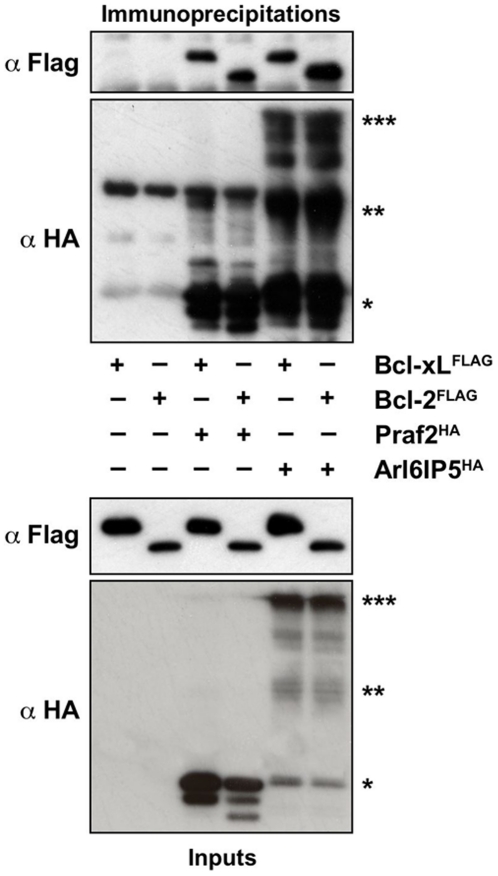
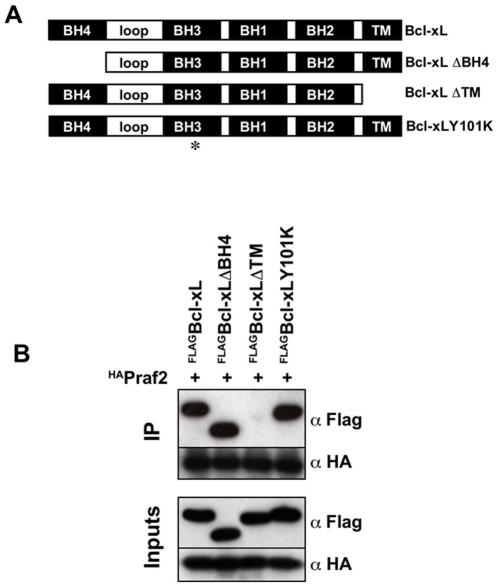
Increased expression of Bcl-xL in cancer has been shown to confer resistance to a broad range of apoptotic stimuli and to modulate a number of other aspects of cellular physiology, including energy metabolism, cell cycle, autophagy, mitochondrial fission/fusion and cellular adhesion. However, only few of these activities have a mechanistic explanation. Here we used Tandem Affinity purification to identify novel Bcl-xL interacting proteins that could explain the pleiotropic effects of Bcl-xL overexpression. Among the several proteins co-purifying with Bcl-xL, we focused on Praf2, a protein with a predicted role in trafficking. The interaction of Praf2 with Bcl-xL was found to be dependent on the transmembrane domain of Bcl-xL. We found that Bcl-2 also interacts with Praf2 and that Bcl-xL and Bcl-2 can interact also with Arl6IP5, an homologue of Praf2. Interestingly, overexpression of Praf2 results in the translocation of Bax to mitochondria and the induction of apoptotic cell death. Praf2 dependent cell death is prevented by the co-transfection of Bcl-xL but not by its transmembrane domain deleted mutant. Accordingly, knock-down of Praf2 increases clonogenicity of U2OS cells following etoposide treatment by reducing cell death. In conclusion a screen for Bcl-xL-interacting membrane proteins let us identify a novel proapoptotic protein whose activity is strongly counteracted exclusively by membrane targeted Bcl-xL.
Conflict of interest statement
Figures




References
-
- Iaccarino I, Hancock D, Evan G, Downward J. c-Myc induces cytochrome c release in Rat1 fibroblasts by increasing outer mitochondrial membrane permeability in a Bid-dependent manner. Cell Death Differ. 2003;10:599–608. - PubMed
-
- Dong Z, Wang J. Hypoxia selection of death-resistant cells. A role for Bcl-X(L). J Biol Chem. 2004;279:9215–9221. - PubMed
-
- Kauffmann-Zeh A, Rodriguez-Viciana P, Ulrich E, Gilbert C, Coffer P, et al. Suppression of c-Myc-induced apoptosis by Ras signalling through PI(3)K and PKB. Nature. 1997;385:544–548. - PubMed
-
- Amundson SA, Myers TG, Scudiero D, Kitada S, Reed JC, et al. An Informatics Approach Identifying Markers of Chemosensitivity in Human Cancer Cell Lines. Cancer Res. 2000;60:6101–6110. - PubMed
-
- Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death. Nat Rev Mol Cell Biol. 2008;9:47–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
