

Prox1 regulates the notch1-mediated inhibition of neurogenesis
- PMID: 21203589
- PMCID: PMC3006385
- DOI: 10.1371/journal.pbio.1000565
Prox1 regulates the notch1-mediated inhibition of neurogenesis
Abstract
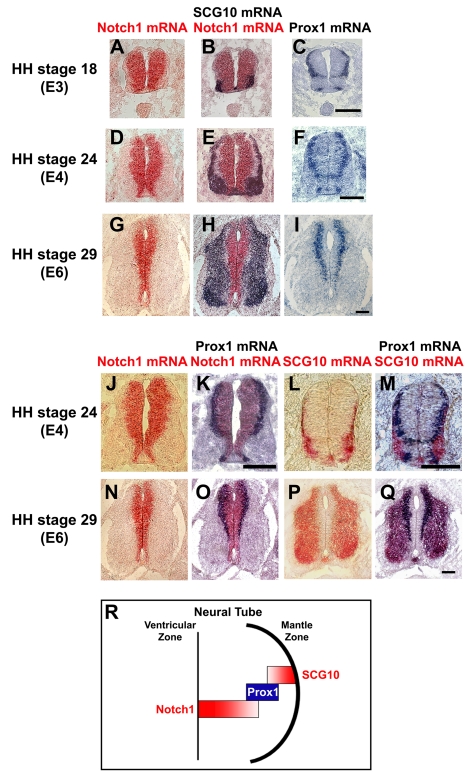
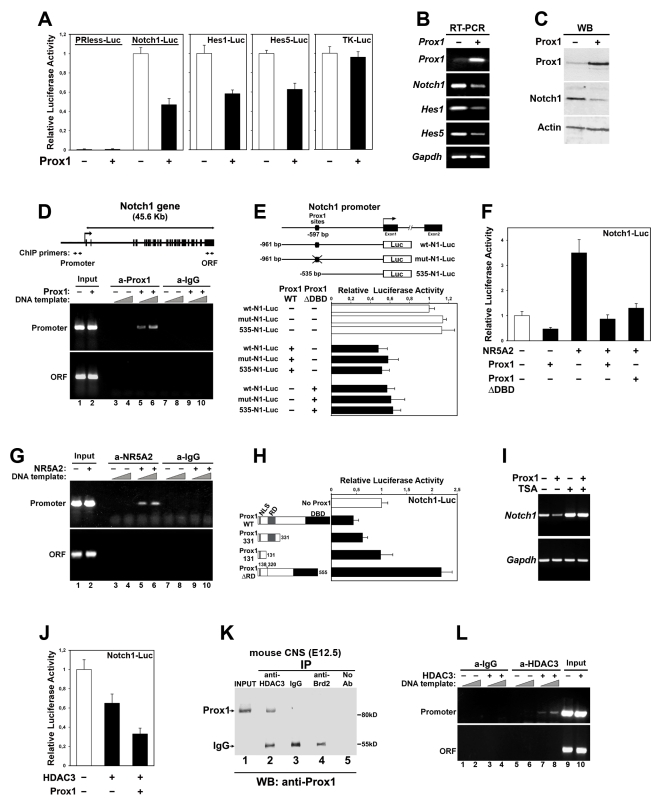
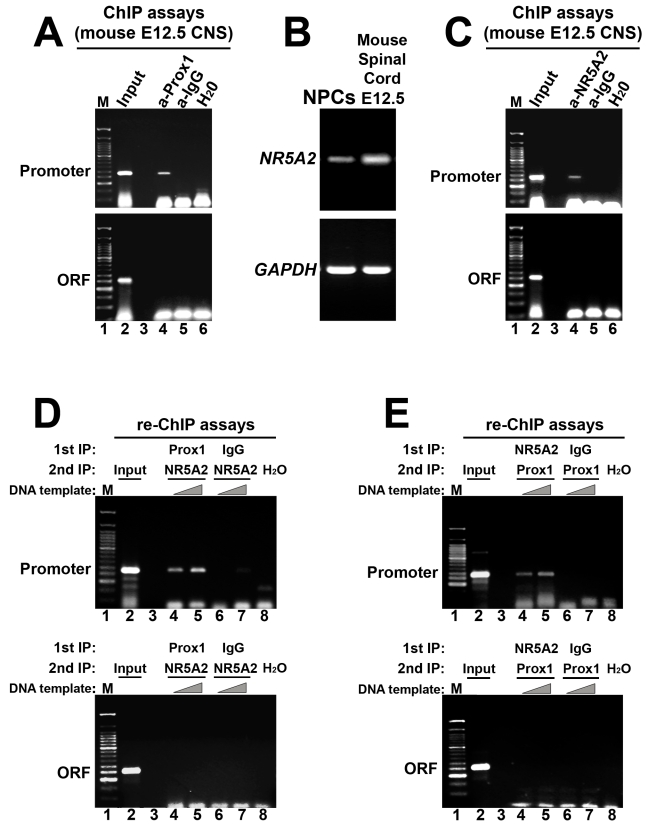
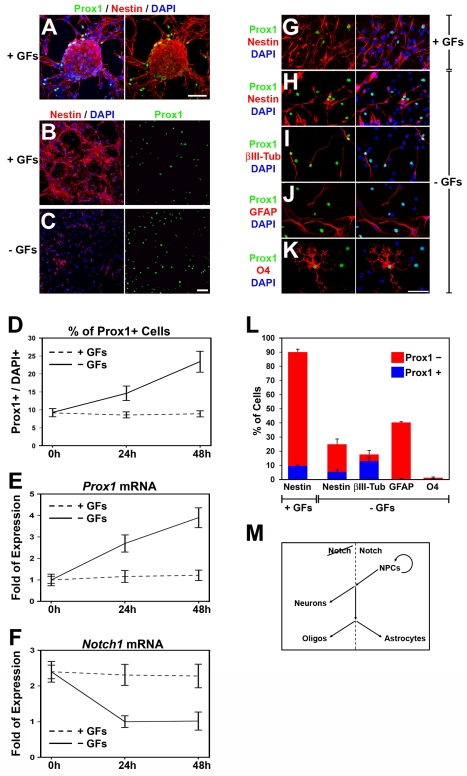
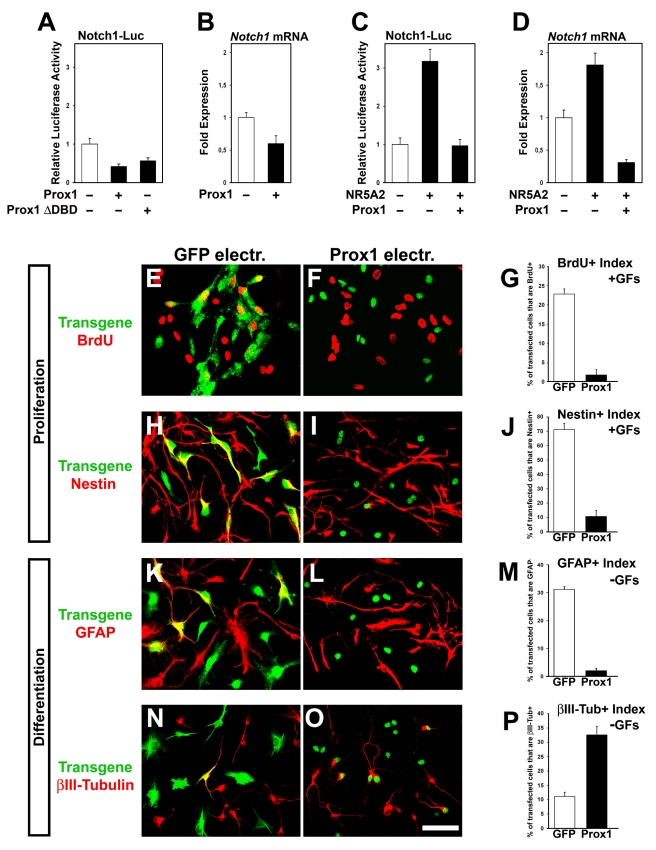
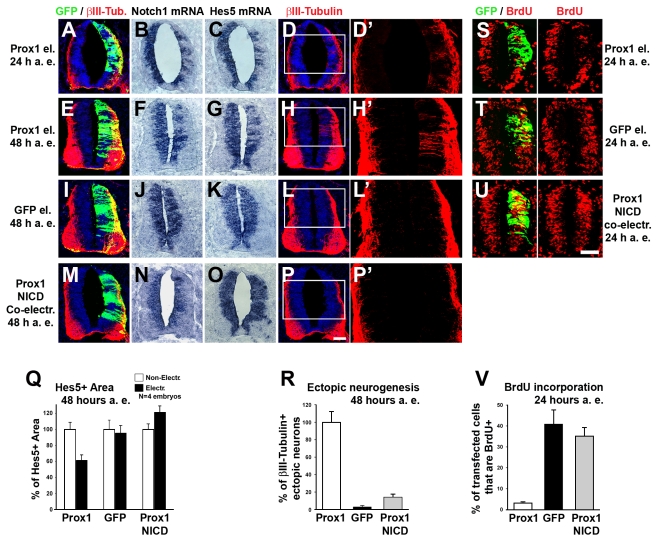
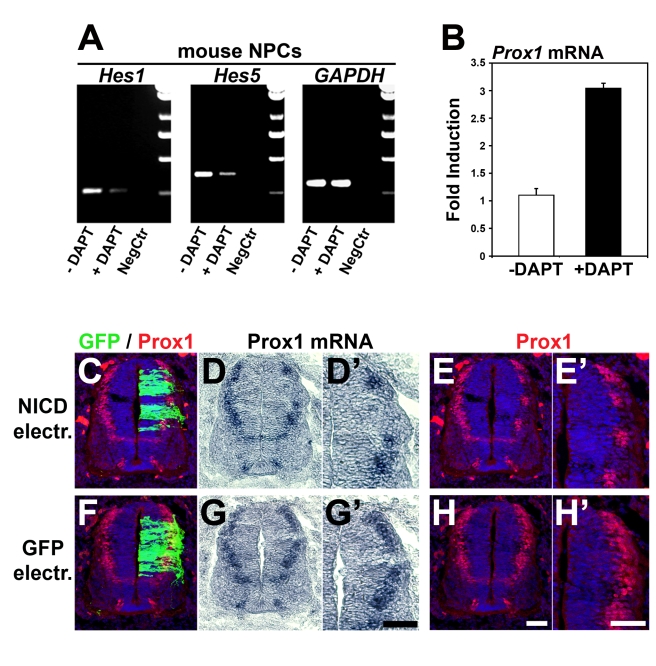
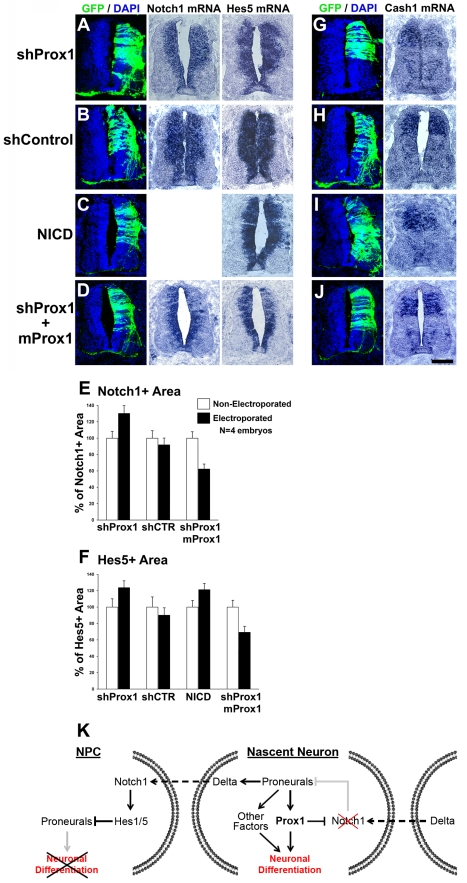
Activation of Notch1 signaling in neural progenitor cells (NPCs) induces self-renewal and inhibits neurogenesis. Upon neuronal differentiation, NPCs overcome this inhibition, express proneural genes to induce Notch ligands, and activate Notch1 in neighboring NPCs. The molecular mechanism that coordinates Notch1 inactivation with initiation of neurogenesis remains elusive. Here, we provide evidence that Prox1, a transcription repressor and downstream target of proneural genes, counteracts Notch1 signaling via direct suppression of Notch1 gene expression. By expression studies in the developing spinal cord of chick and mouse embryo, we showed that Prox1 is limited to neuronal precursors residing between the Notch1+ NPCs and post-mitotic neurons. Physiological levels of Prox1 in this tissue are sufficient to allow binding at Notch1 promoter and they are critical for proper Notch1 transcriptional regulation in vivo. Gain-of-function studies in the chick neural tube and mouse NPCs suggest that Prox1-mediated suppression of Notch1 relieves its inhibition on neurogenesis and allows NPCs to exit the cell cycle and differentiate. Moreover, loss-of-function in the chick neural tube shows that Prox1 is necessary for suppression of Notch1 outside the ventricular zone, inhibition of active Notch signaling, down-regulation of NPC markers, and completion of neuronal differentiation program. Together these data suggest that Prox1 inhibits Notch1 gene expression to control the balance between NPC self-renewal and neuronal differentiation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Kageyama R, Ohtsuka T, Shimojo H, Imayoshi I. Dynamic Notch signaling in neural progenitor cells and a revised view of lateral inhibition. Nat Neurosci. 2008;11:1247–1251. - PubMed
-
- Louvi A, Artavanis-Tsakonas S. Notch signalling in vertebrate neural development. Nat Rev Neurosci. 2006;7:93–102. - PubMed
-
- Yoon K, Gaiano N. Notch signaling in the mammalian central nervous system: insights from mouse mutants. Nat Neurosci. 2005;8:709–715. - PubMed
-
- Selkoe D, Kopan R. Notch and Presenilin: regulated intramembrane proteolysis links development and degeneration. Annu Rev Neurosci. 2003;26:565–597. - PubMed
-
- Bertrand N, Castro D. S, Guillemot F. Proneural genes and the specification of neural cell types. Nat Rev Neurosci. 2002;3:517–530. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
