

Peptidomic analysis of human cell lines
- PMID: 21204522
- PMCID: PMC3070057
- DOI: 10.1021/pr100952f
Peptidomic analysis of human cell lines
Abstract
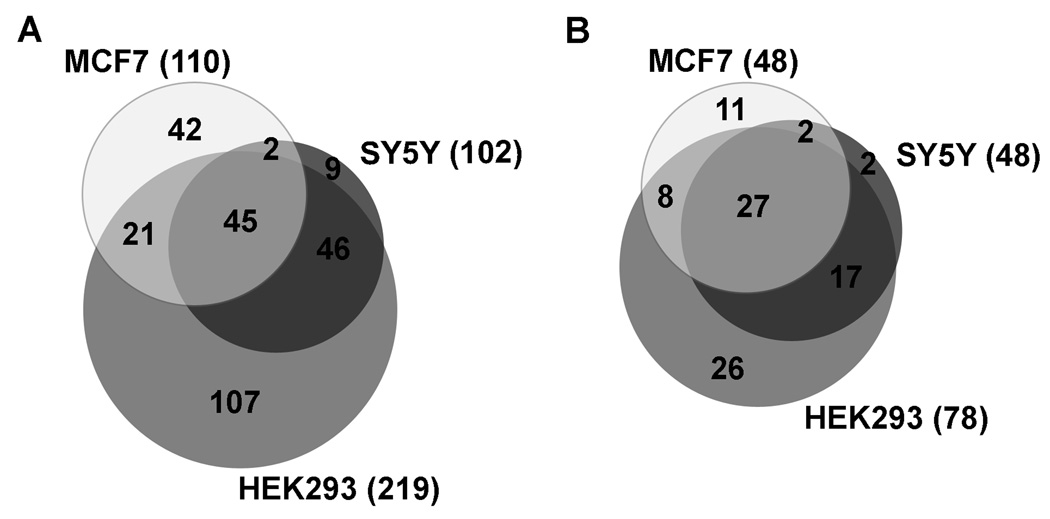
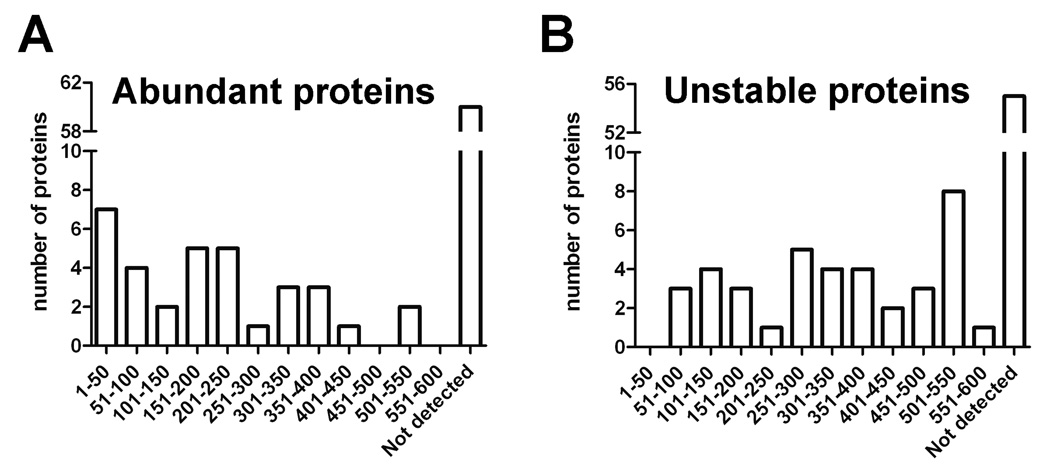
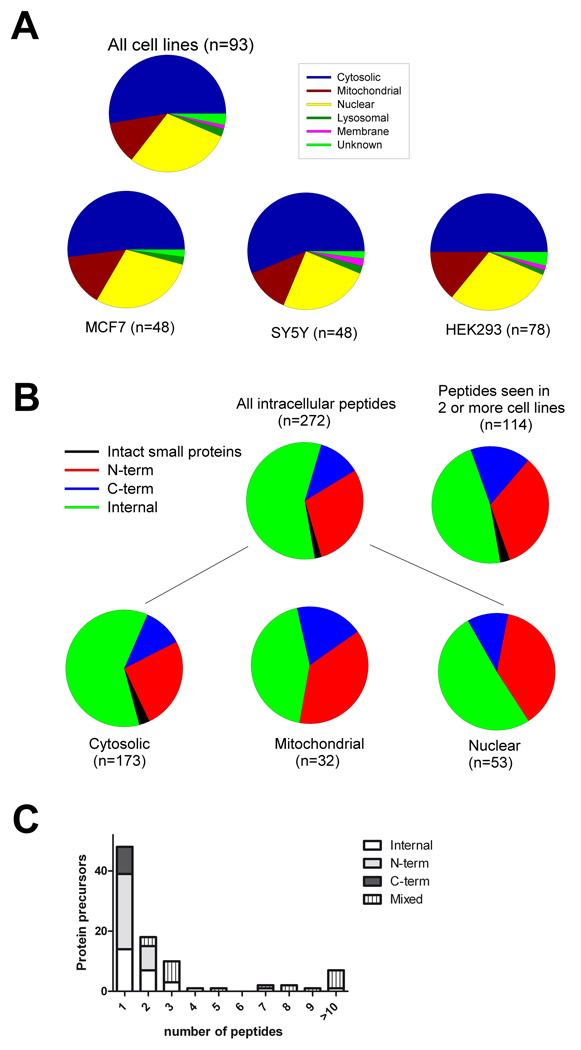
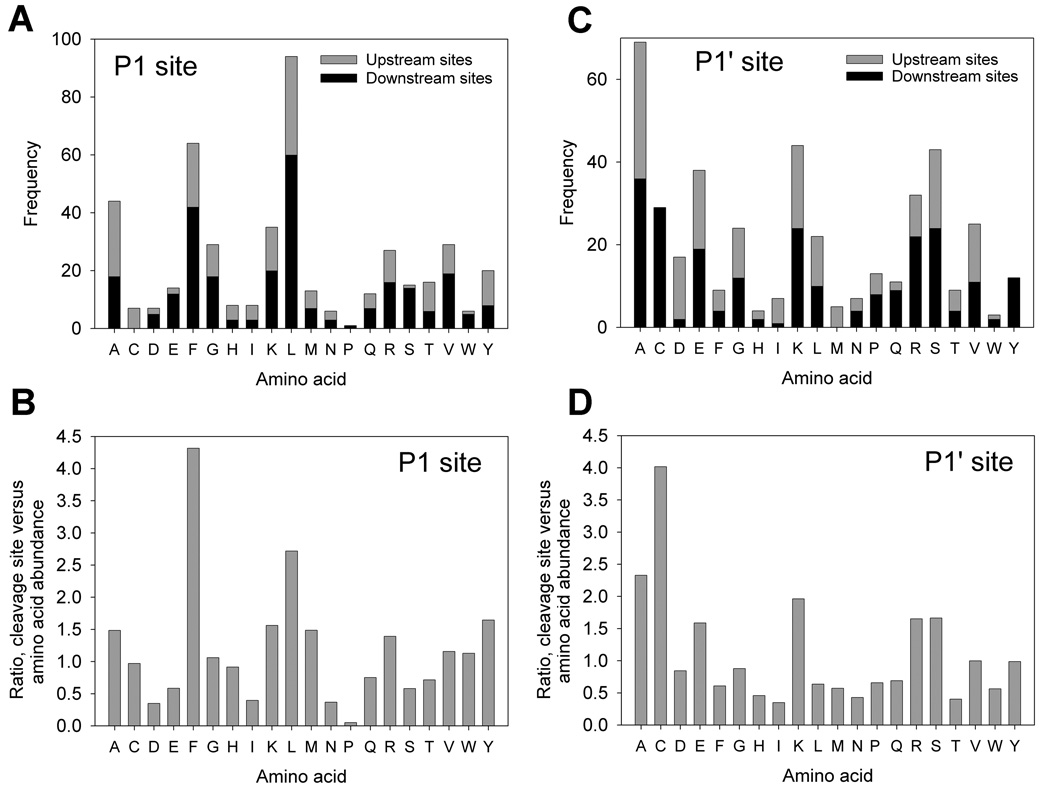
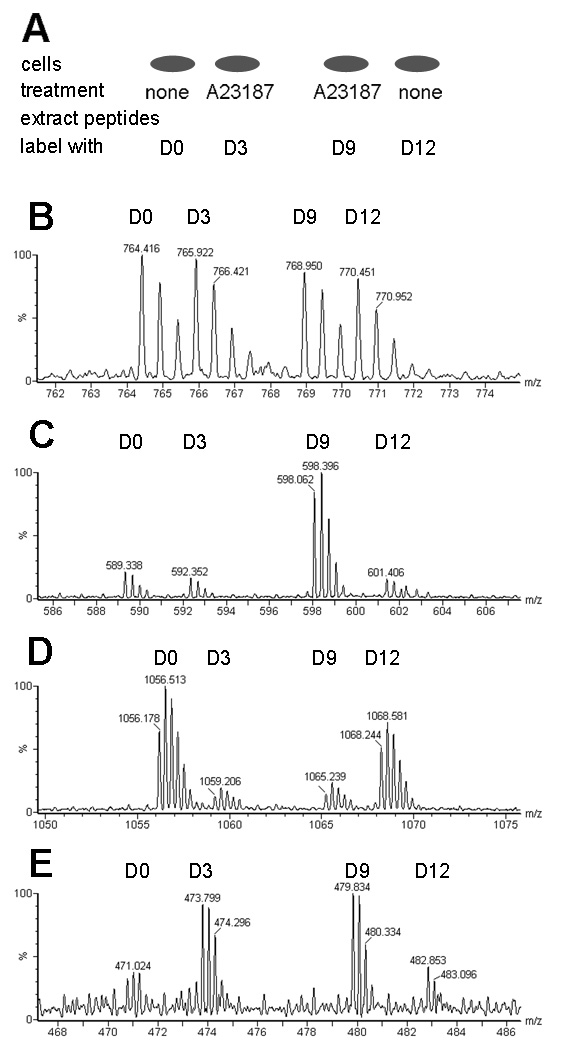
Peptides have been proposed to function in intracellular signaling within the cytosol. Although cytosolic peptides are considered to be highly unstable, a large number of peptides have been detected in mouse brain and other biological samples. In the present study, we evaluated the peptidome of three diverse cell lines: SH-SY5Y, MCF7, and HEK293 cells. A comparison of the peptidomes revealed considerable overlap in the identity of the peptides found in each cell line. The majority of the observed peptides are not derived from the most abundant or least stable proteins in the cell, and approximately half of the cellular peptides correspond to the N- or C- termini of the precursor proteins. Cleavage site analysis revealed a preference for hydrophobic residues in the P1 position. Quantitative peptidomic analysis indicated that the levels of most cellular peptides are not altered in response to elevated intracellular calcium, suggesting that calpain is not responsible for their production. The similarity of the peptidomes of the three cell lines and the lack of correlation with the predicted cellular degradome implies the selective formation or retention of these peptides, consistent with the hypothesis that they are functional in the cells.
Figures
References
-
- Wysocki VH, Resing KA, Zhang Q, Cheng G. Methods. 2005;35:211–222. - PubMed
-
- Fricker LD, Lim J, Pan H, Che FY. Mass Spectrom Rev. 2006;25:327–344. - PubMed
-
- Svensson M, Skold K, Nilsson A, Falth M, Svenningsson P, Andren PE. Biochem Soc Trans. 2007;35:588–593. - PubMed
-
- Cape SS, Dowell JA, Li L. Methods Mol Biol. 2009;492:381–393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
