

Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice
- PMID: 21206087
- PMCID: PMC3026727
- DOI: 10.1172/JCI43984
Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice
Erratum in
-
Vaccine-induced protection against 3 systemic mycoses endemic to North America requires Th17 cells in mice.J Clin Invest. 2016 Feb;126(2):795. doi: 10.1172/JCI85788. Epub 2016 Feb 1. J Clin Invest. 2016. PMID: 26829626 Free PMC article. No abstract available.
Abstract
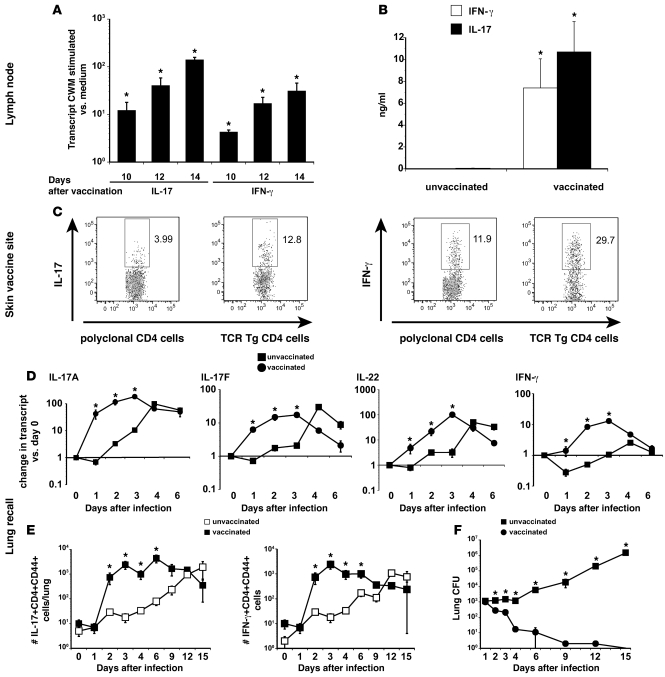
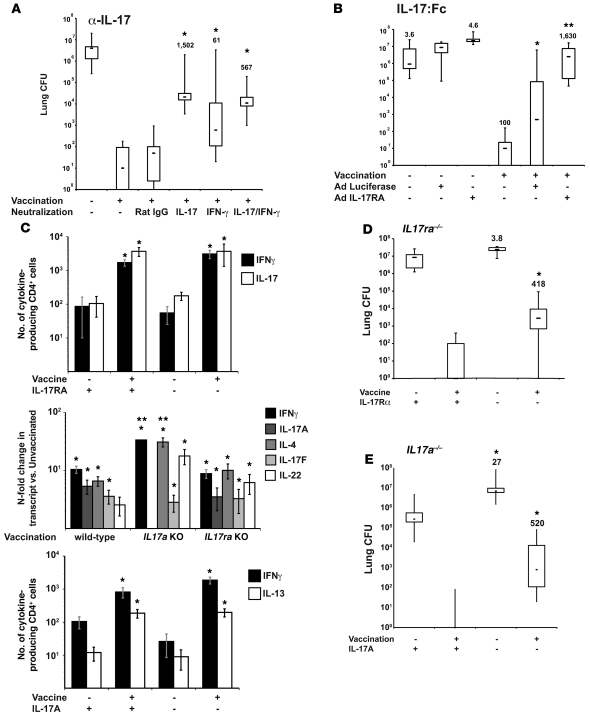
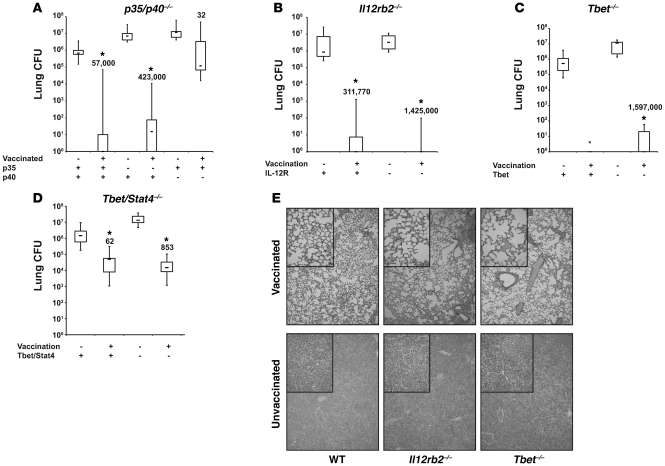
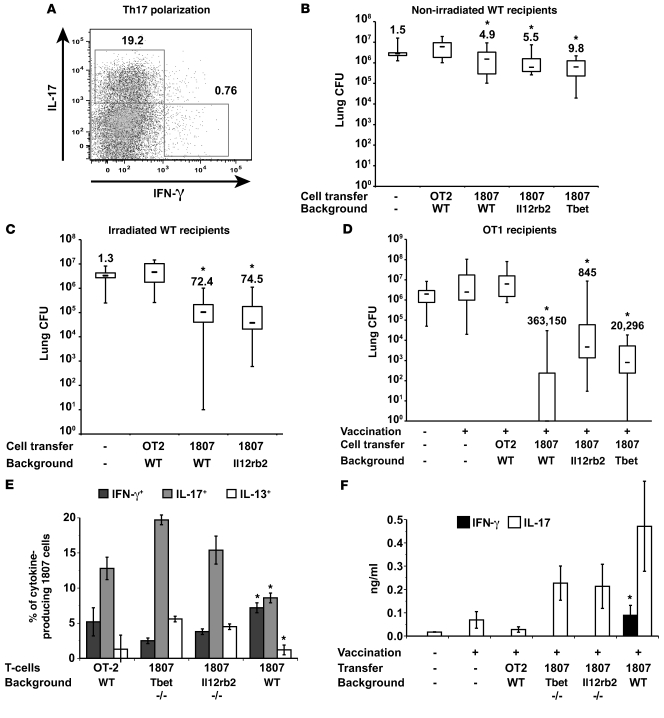
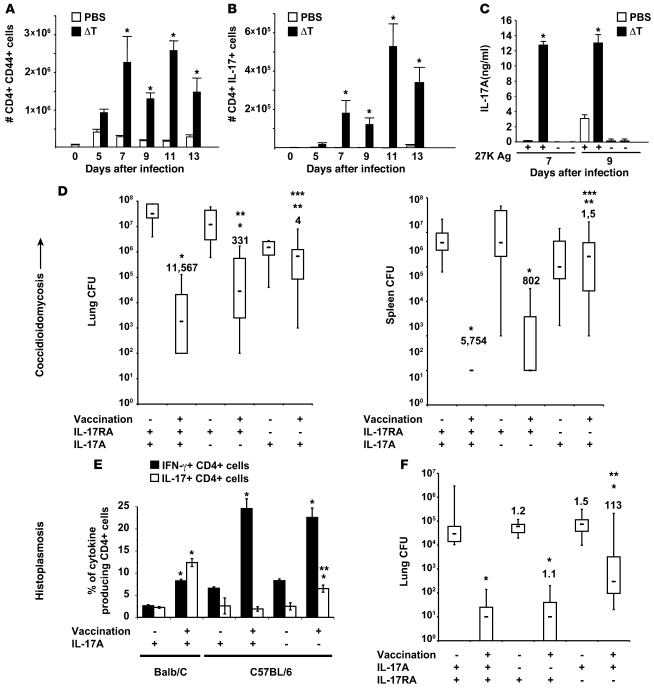
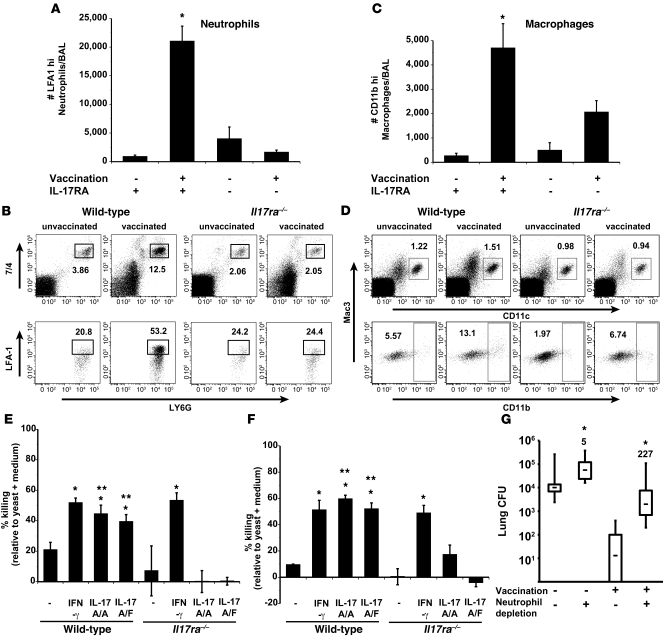
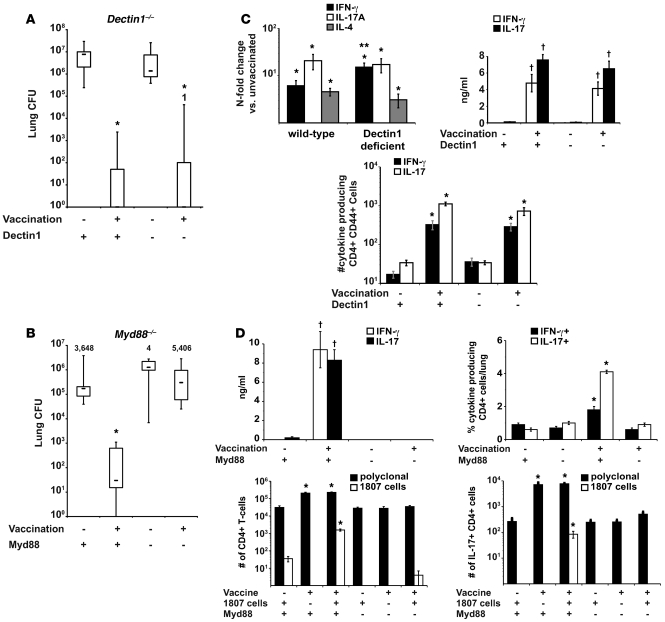
Worldwide rates of systemic fungal infections, including three of the major pathogens responsible for such infections in North America (Coccidioides posadasii, Histoplasma capsulatum, and Blastomyces dermatitidis), have soared recently, spurring interest in developing vaccines. The development of Th1 cells is believed to be crucial for protective immunity against pathogenic fungi, whereas the role of Th17 cells is vigorously debated. In models of primary fungal infection, some studies have shown that Th17 cells mediate resistance, while others have shown that they promote disease pathology. Here, we have shown that Th1 immunity is dispensable and that fungus-specific Th17 cells are sufficient for vaccine-induced protection against lethal pulmonary infection with B. dermatitidis in mice. Further, vaccine-induced Th17 cells were necessary and sufficient to protect against the three major systemic mycoses in North America. Mechanistically, Th17 cells engendered protection by recruiting and activating neutrophils and macrophages to the alveolar space, while the induction of Th17 cells and acquisition of vaccine immunity unexpectedly required the adapter molecule Myd88 but not the fungal pathogen recognition receptor Dectin-1. These data suggest that human vaccines against systemic fungal infections should be designed to induce Th17 cells if they are to be effective.
Figures
Comment in
-
Vaccines: Foes of fungi are just 17.Nat Rev Immunol. 2011 Feb;11(2):74. doi: 10.1038/nri2929. Nat Rev Immunol. 2011. PMID: 21467978 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
