

Peak frequency in the theta and alpha bands correlates with human working memory capacity
- PMID: 21206531
- PMCID: PMC3009479
- DOI: 10.3389/fnhum.2010.00200
Peak frequency in the theta and alpha bands correlates with human working memory capacity
Abstract
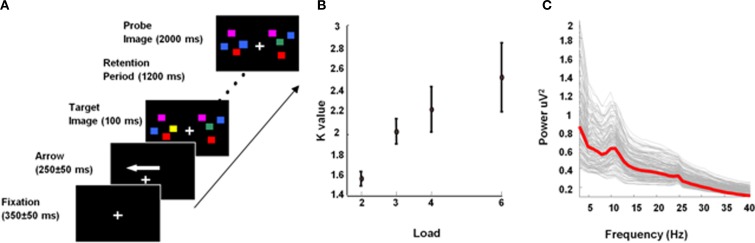
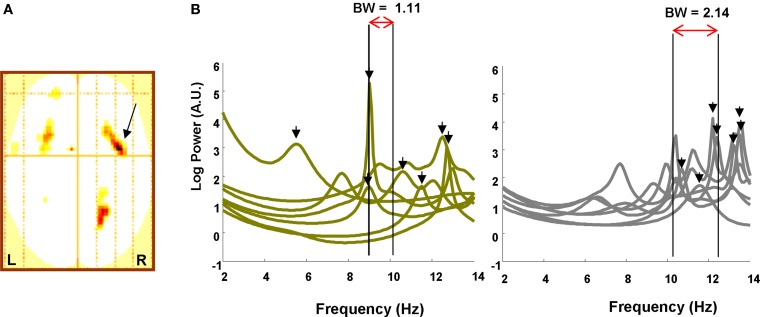
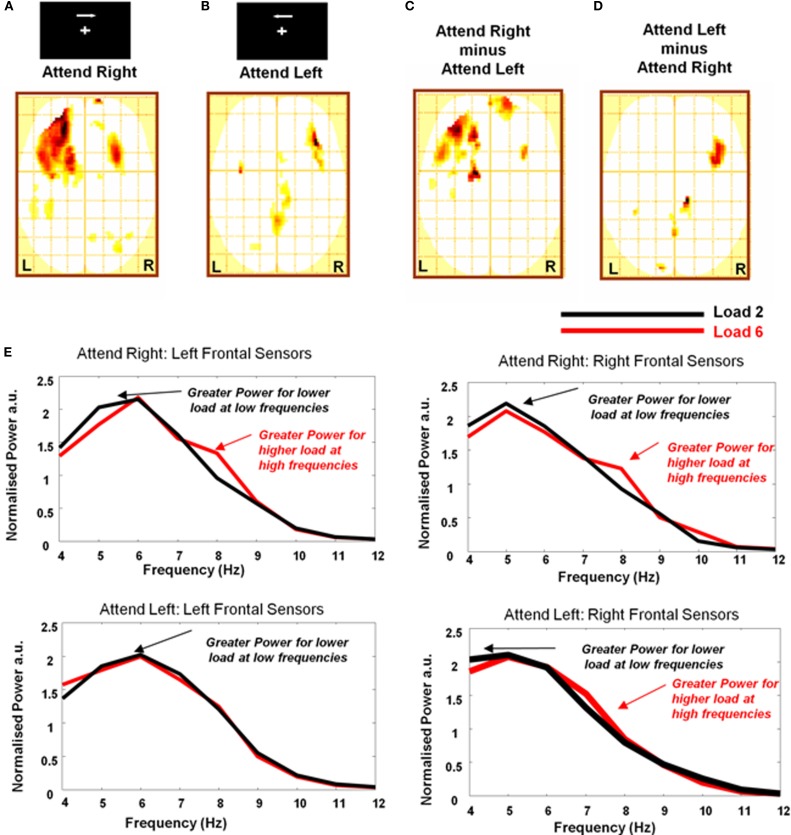
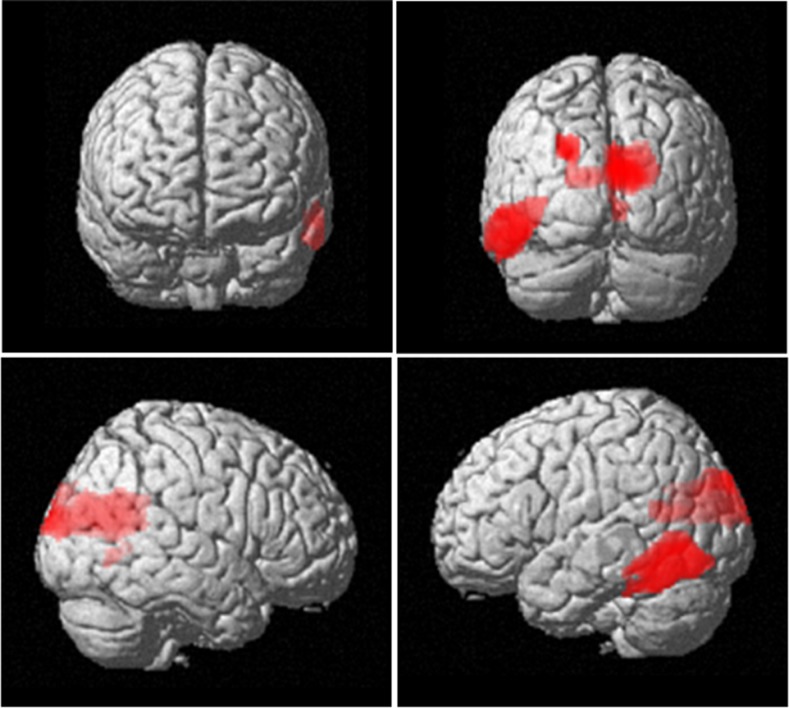
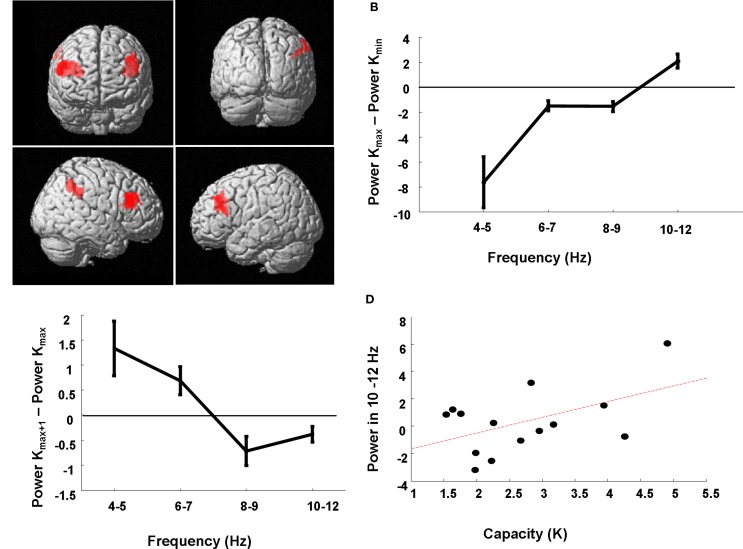
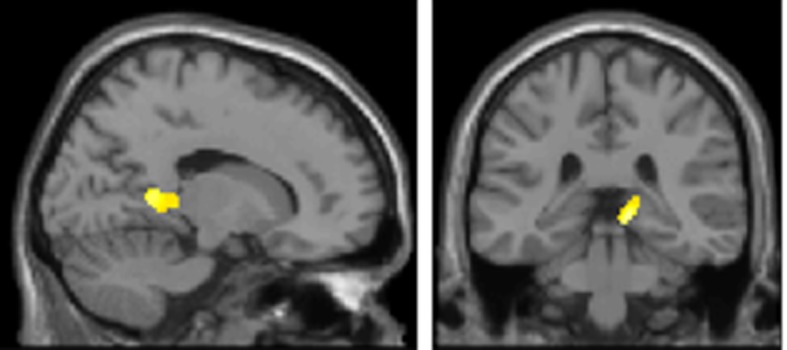
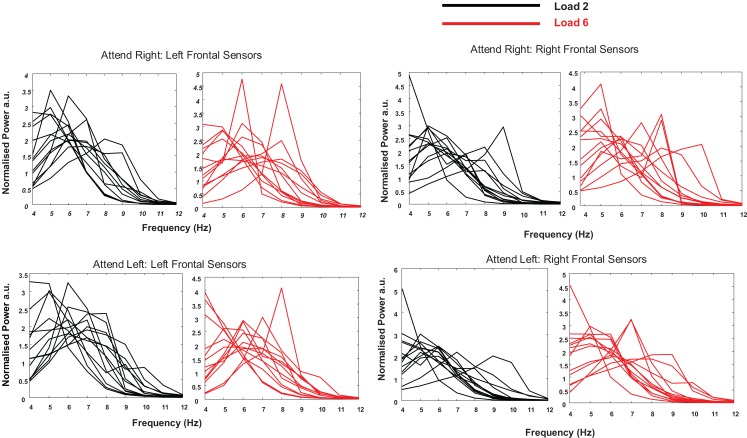
Theta oscillations in the local field potential of neural ensembles are considered key mediators of human working memory. Theoretical accounts arising from animal hippocampal recordings propose that the phase of theta oscillations serves to instantiate sequential neuronal firing to form discrete representations of items held online. Human evidence of phase relationships in visual working memory has enhanced this theory, implicating long theta cycles in supporting greater memory capacity. Here we use human magnetoencephalographic recordings to examine a novel, alternative principle of theta functionality. The principle we hypothesize is derived from information theory and predicts that rather than long (low frequency) theta cycles, short (high frequency) theta cycles are best suited to support high information capacity. From oscillatory activity recorded during the maintenance period of a visual working memory task we show that a network of brain regions displays an increase in peak 4-12 Hz frequency with increasing memory load. Source localization techniques reveal that this network comprises bilateral prefrontal and right parietal cortices. Further, the peak of oscillation along this theta-alpha frequency axis is significantly higher in high capacity individuals compared to low capacity individuals. Importantly while we observe the adherence of cortical neuronal oscillations to our novel principle of theta functioning, we also observe the traditional inverse effect of low frequency theta maintaining high loads, where critically this was located in medial temporal regions suggesting parallel, dissociable hippocampal-centric, and prefrontal-centric theta mechanisms.
Keywords: MEG; alpha; capacity; oscillations; theta; visual working memory.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources