

Adenine nucleotide transporters in organelles: novel genes and functions
- PMID: 21207102
- PMCID: PMC11114886
- DOI: 10.1007/s00018-010-0612-3
Adenine nucleotide transporters in organelles: novel genes and functions
Abstract
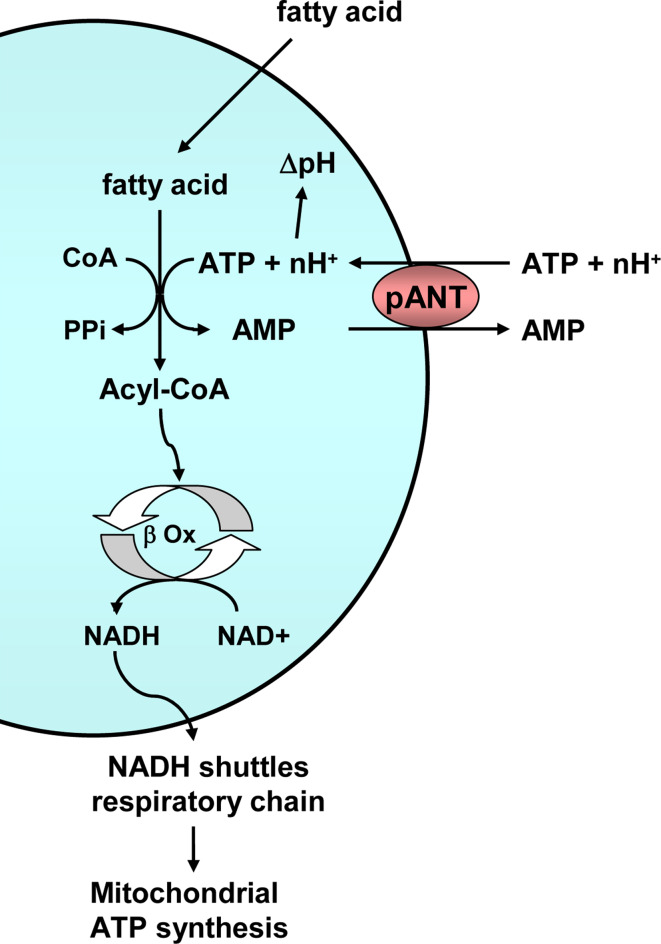
In eukaryotes, cellular energy in the form of ATP is produced in the cytosol via glycolysis or in the mitochondria via oxidative phosphorylation and, in photosynthetic organisms, in the chloroplast via photophosphorylation. Transport of adenine nucleotides among cell compartments is essential and is performed mainly by members of the mitochondrial carrier family, among which the ADP/ATP carriers are the best known. This work reviews the carriers that transport adenine nucleotides into the organelles of eukaryotic cells together with their possible functions. We focus on novel mechanisms of adenine nucleotide transport, including mitochondrial carriers found in organelles such as peroxisomes, plastids, or endoplasmic reticulum and also mitochondrial carriers found in the mitochondrial remnants of many eukaryotic parasites of interest. The extensive repertoire of adenine nucleotide carriers highlights an amazing variety of new possible functions of adenine nucleotide transport across eukaryotic organelles.
Figures
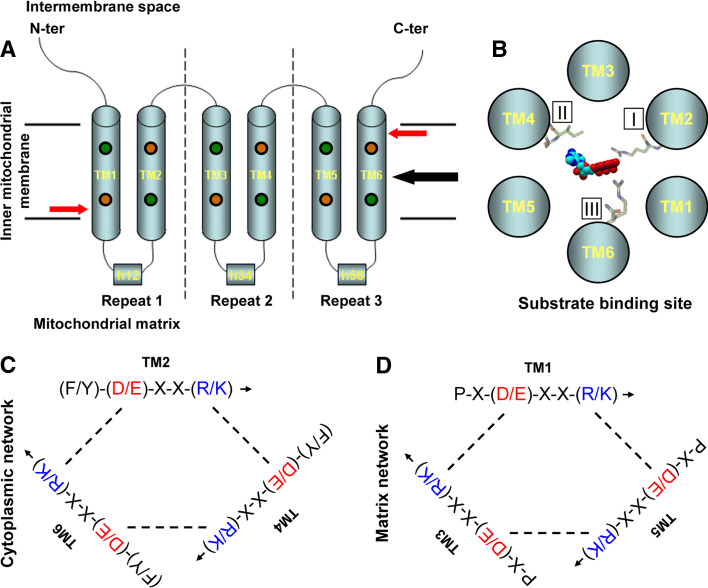
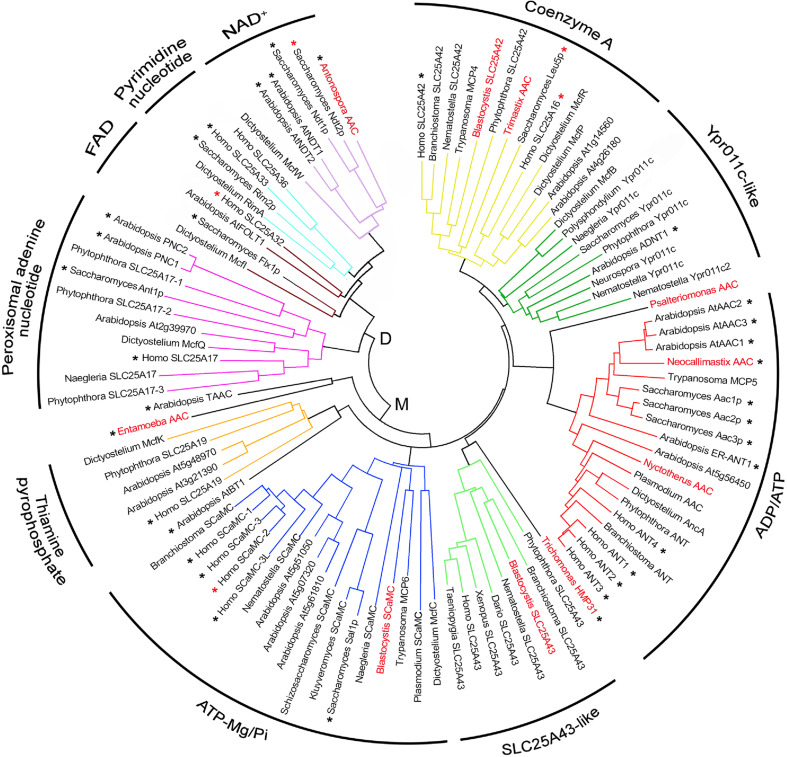
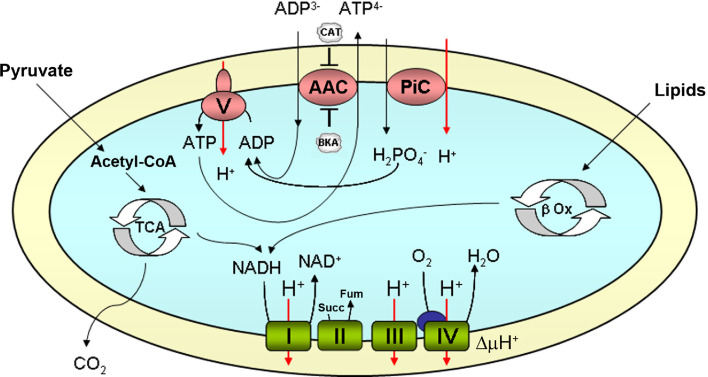
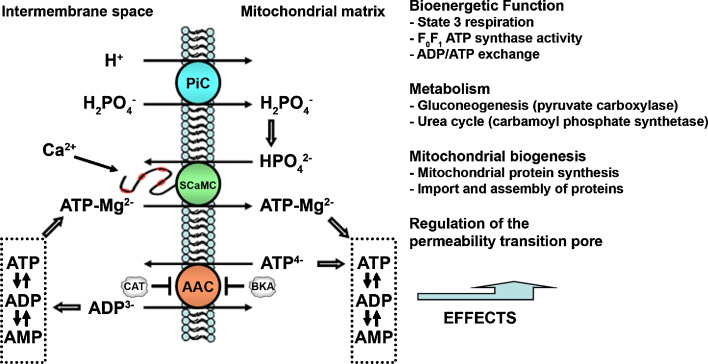
Similar articles
-
A novel ADP/ATP transporter in the mitosome of the microaerophilic human parasite Entamoeba histolytica.Curr Biol. 2005 Apr 26;15(8):737-42. doi: 10.1016/j.cub.2005.02.068. Curr Biol. 2005. PMID: 15854906
-
The ADP and ATP transport in mitochondria and its carrier.Biochim Biophys Acta. 2008 Oct;1778(10):1978-2021. doi: 10.1016/j.bbamem.2008.04.011. Epub 2008 May 2. Biochim Biophys Acta. 2008. PMID: 18510943 Review.
-
A Single Cysteine Residue in the Translocation Pathway of the Mitosomal ADP/ATP Carrier from Cryptosporidium parvum Confers a Broad Nucleotide Specificity.Int J Mol Sci. 2020 Nov 26;21(23):8971. doi: 10.3390/ijms21238971. Int J Mol Sci. 2020. PMID: 33255957 Free PMC article.
-
A divergent ADP/ATP carrier in the hydrogenosomes of Trichomonas gallinae argues for an independent origin of these organelles.Mol Microbiol. 2004 Mar;51(5):1439-46. doi: 10.1111/j.1365-2958.2004.03918.x. Mol Microbiol. 2004. PMID: 14982636
-
Transport of adenine nucleotides in the mitochondria of Saccharomyces cerevisiae: interactions between the ADP/ATP carriers and the ATP-Mg/Pi carrier.Mitochondrion. 2009 Apr;9(2):79-85. doi: 10.1016/j.mito.2009.01.001. Epub 2009 Jan 17. Mitochondrion. 2009. PMID: 19460304 Review.
Cited by
-
Mitonuclear conflict and cooperation govern the integration of genotypes, phenotypes and environments.Philos Trans R Soc Lond B Biol Sci. 2020 Jan 20;375(1790):20190188. doi: 10.1098/rstb.2019.0188. Epub 2019 Dec 2. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 31787039 Free PMC article.
-
The plant mitochondrial carrier family: functional and evolutionary aspects.Front Plant Sci. 2012 Jan 18;3:2. doi: 10.3389/fpls.2012.00002. eCollection 2012. Front Plant Sci. 2012. PMID: 22639632 Free PMC article.
-
Physiological and pathological roles of mitochondrial SLC25 carriers.Biochem J. 2013 Sep 15;454(3):371-86. doi: 10.1042/BJ20121753. Biochem J. 2013. PMID: 23988125 Free PMC article. Review.
-
A self-sequestered calmodulin-like Ca²⁺ sensor of mitochondrial SCaMC carrier and its implication to Ca²⁺-dependent ATP-Mg/P(i) transport.Structure. 2014 Feb 4;22(2):209-17. doi: 10.1016/j.str.2013.10.018. Epub 2013 Dec 12. Structure. 2014. PMID: 24332718 Free PMC article.
-
De Novo Mutations in SLC25A24 Cause a Disorder Characterized by Early Aging, Bone Dysplasia, Characteristic Face, and Early Demise.Am J Hum Genet. 2017 Nov 2;101(5):844-855. doi: 10.1016/j.ajhg.2017.09.017. Am J Hum Genet. 2017. PMID: 29100094 Free PMC article.
References
-
- Palmieri F, Agrimi G, Blanco E, Castegna A, Di Noia MA, Iacobazzi V, Lasorsa FM, Marobbio CM, Palmieri L, Scarcia P, Todisco S, Vozza A, Walker J. Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim Biophys Acta. 2006;1757:1249–1262. doi: 10.1016/j.bbabio.2006.05.023. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
