

Although divergent in residues of the peptide binding site, conserved chimpanzee Patr-AL and polymorphic human HLA-A*02 have overlapping peptide-binding repertoires
- PMID: 21209280
- PMCID: PMC3124313
- DOI: 10.4049/jimmunol.1002990
Although divergent in residues of the peptide binding site, conserved chimpanzee Patr-AL and polymorphic human HLA-A*02 have overlapping peptide-binding repertoires
Abstract
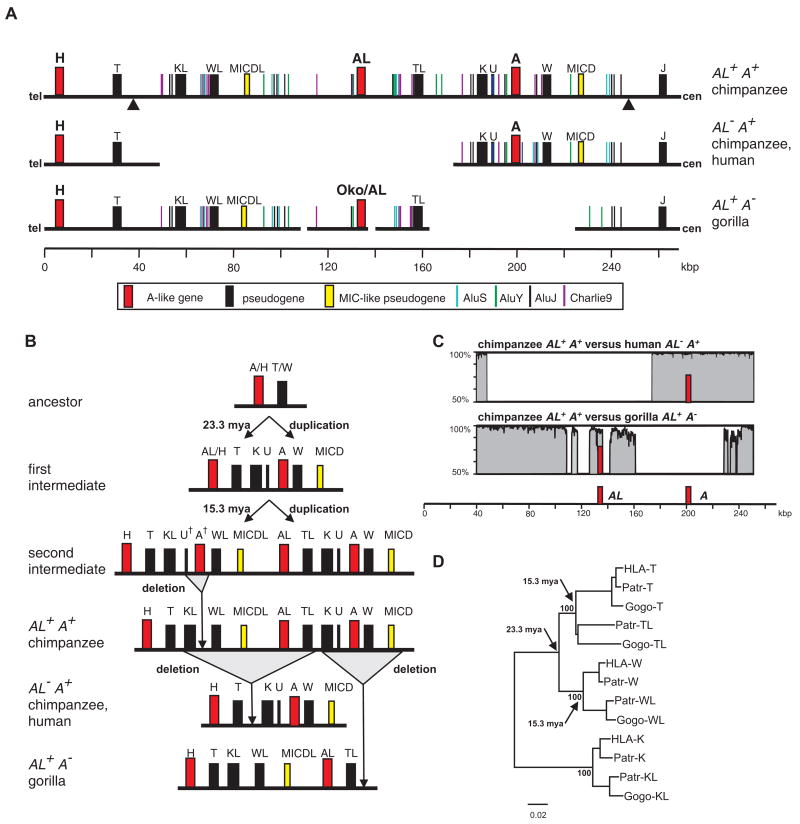
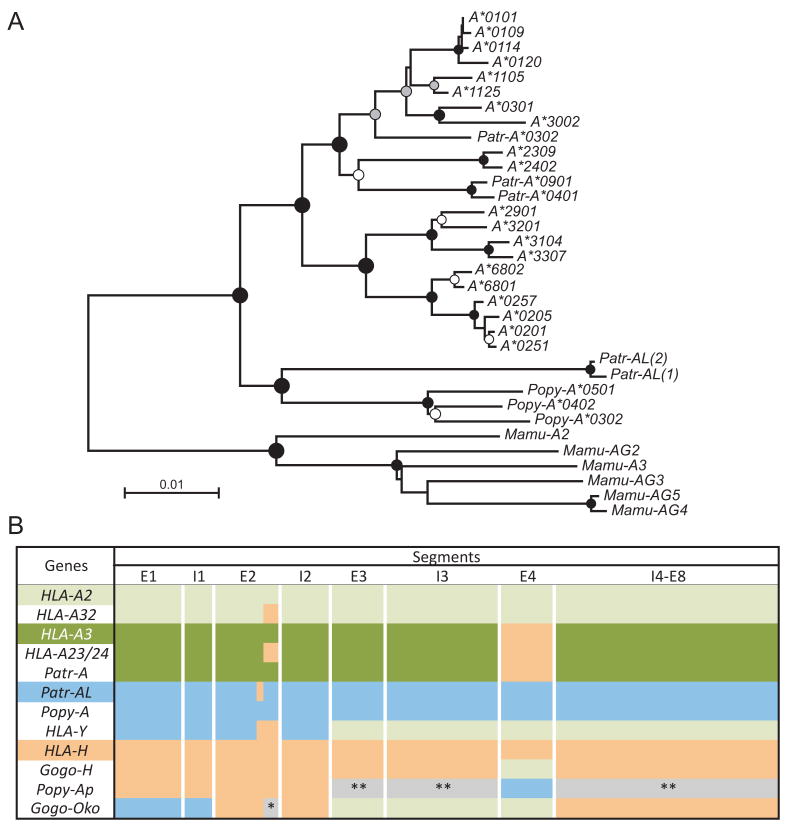
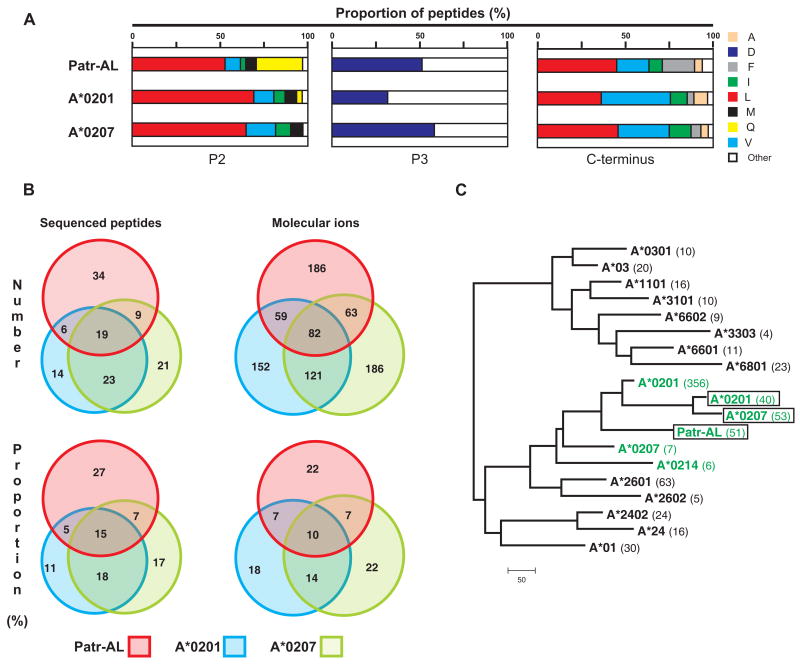
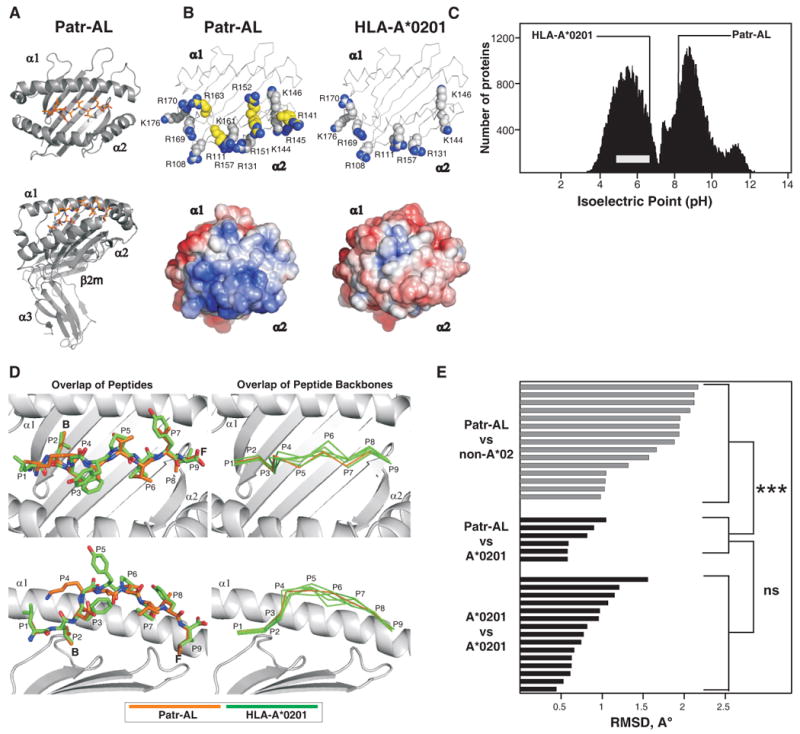
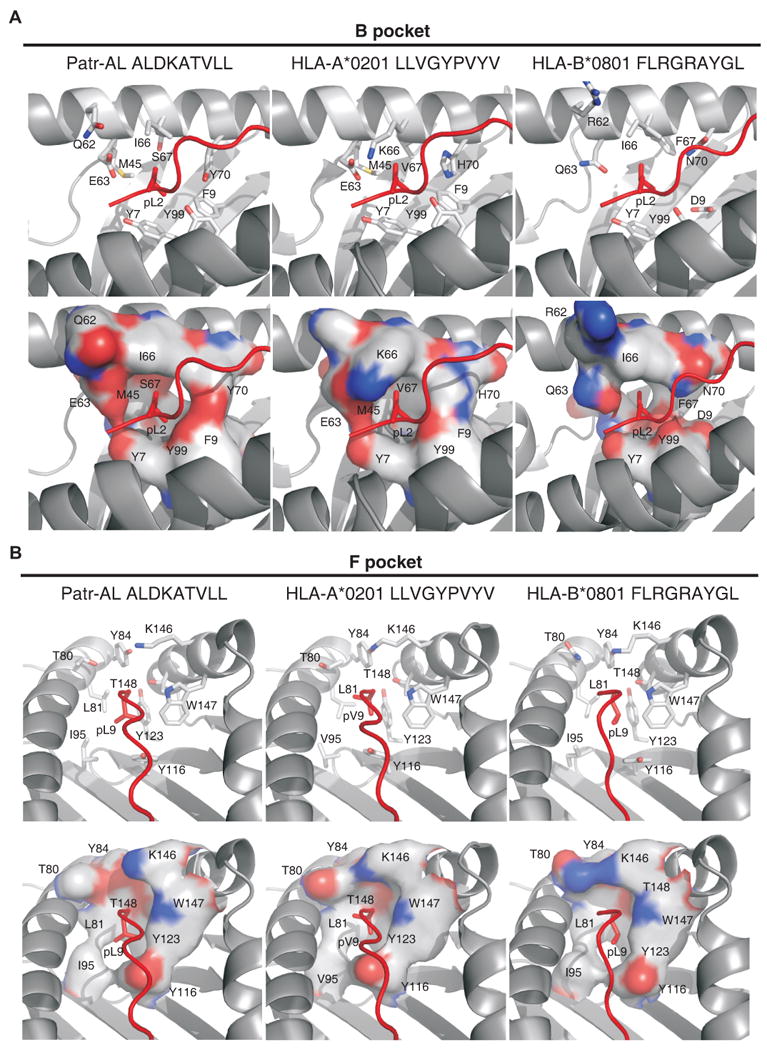
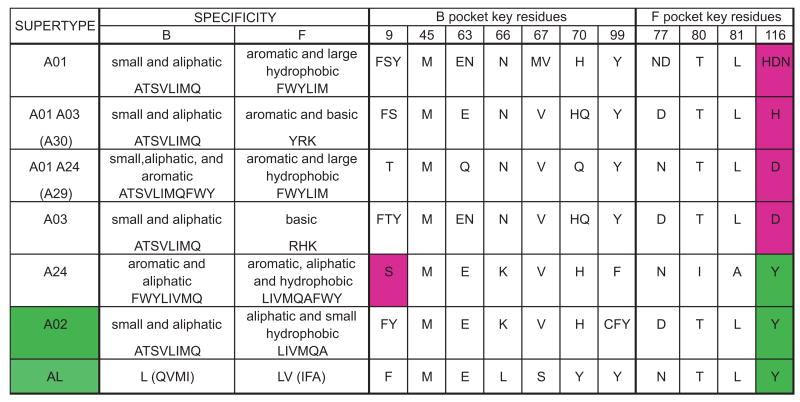
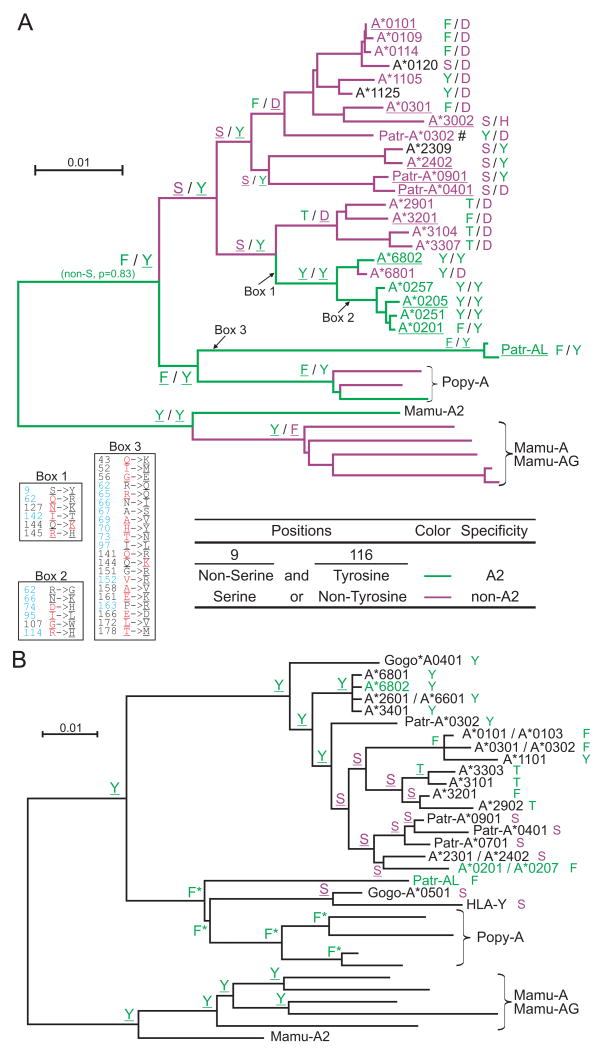
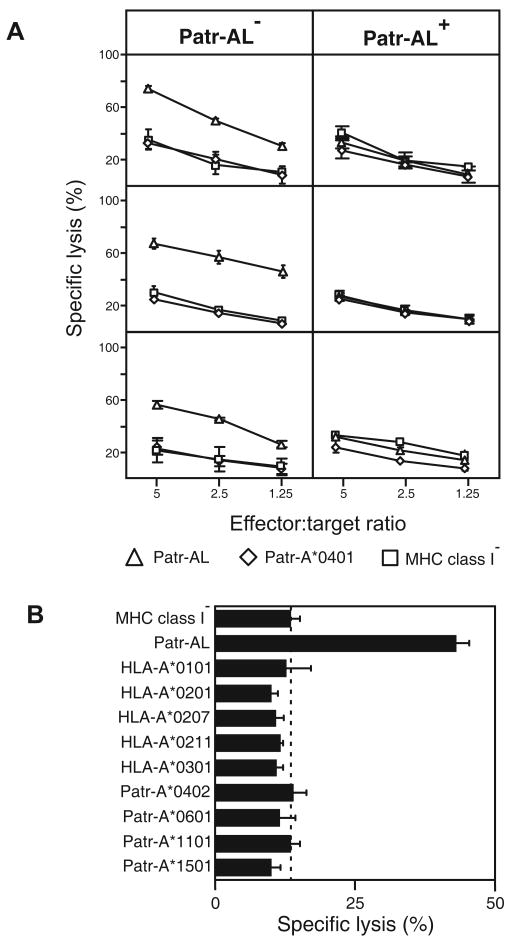
Patr-AL is an expressed, non-polymorphic MHC class I gene carried by ∼50% of chimpanzee MHC haplotypes. Comparing Patr-AL(+) and Patr-AL(-) haplotypes showed Patr-AL defines a unique 125-kb genomic block flanked by blocks containing classical Patr-A and pseudogene Patr-H. Orthologous to Patr-AL are polymorphic orangutan Popy-A and the 5' part of human pseudogene HLA-Y, carried by ∼10% of HLA haplotypes. Thus, the AL gene alternatively evolved in these closely related species to become classical, nonclassical, and nonfunctional. Although differing by 30 aa substitutions in the peptide-binding α(1) and α(2) domains, Patr-AL and HLA-A*0201 bind overlapping repertoires of peptides; the overlap being comparable with that between the A*0201 and A*0207 subtypes differing by one substitution. Patr-AL thus has the A02 supertypic peptide-binding specificity. Patr-AL and HLA-A*0201 have similar three-dimensional structures, binding peptides in similar conformation. Although comparable in size and shape, the B and F specificity pockets of Patr-AL and HLA-A*0201 differ in both their constituent residues and contacts with peptide anchors. Uniquely shared by Patr-AL, HLA-A*0201, and other members of the A02 supertype are the absence of serine at position 9 in the B pocket and the presence of tyrosine at position 116 in the F pocket. Distinguishing Patr-AL from HLA-A*02 is an unusually electropositive upper face on the α(2) helix. Stimulating PBMCs from Patr-AL(-) chimpanzees with B cells expressing Patr-AL produced potent alloreactive CD8 T cells with specificity for Patr-AL and no cross-reactivity toward other MHC class I molecules, including HLA-A*02. In contrast, PBMCs from Patr-AL(+) chimpanzees are tolerant of Patr-AL.
Figures
References
-
- Peaper DR, Cresswell P. Regulation of MHC class I assembly and peptide binding. Annu Rev Cell Dev Biol. 2008;24:343–368. - PubMed
-
- Jenkins MK, Chu HH, McLachlan JB, Moon JJ. On the composition of the preimmune repertoire of T cells specific for Peptide-major histocompatibility complex ligands. Annu Rev Immunol. 2010;28:275–294. - PubMed
-
- Adams EJ, Parham P. Species-specific evolution of MHC class I genes in the higher primates. Immunol Rev. 2001;183:41–64. - PubMed
-
- Shiina T, Ota M, Shimizu S, Katsuyama Y, Hashimoto N, Takasu M, Anzai T, Kulski JK, Kikkawa E, Naruse T, Kimura N, Yanagiya K, Watanabe A, Hosomichi K, Kohara S, Iwamoto C, Umehara Y, Meyer A, Wanner V, Sano K, Macquin C, Ikeo K, Tokunaga K, Gojobori T, Inoko H, Bahram S. Rapid evolution of major histocompatibility complex class I genes in primates generates new disease alleles in humans via hitchhiking diversity. Genetics. 2006;173:1555–1570. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
