

Ca2+-dependent structural rearrangements within Na+-Ca2+ exchanger dimers
- PMID: 21209335
- PMCID: PMC3029690
- DOI: 10.1073/pnas.1016114108
Ca2+-dependent structural rearrangements within Na+-Ca2+ exchanger dimers
Abstract
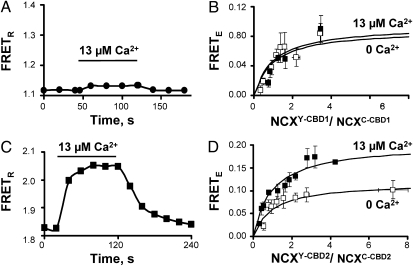
Cytoplasmic Ca(2+) is known to regulate Na(+)-Ca(2+) exchanger (NCX) activity by binding to two adjacent Ca(2+)-binding domains (CBD1 and CBD2) located in the large intracellular loop between transmembrane segments 5 and 6. We investigated Ca(2+)-dependent movements as changes in FRET between exchanger proteins tagged with CFP or YFP at position 266 within the large cytoplasmic loop. Data indicate that the exchanger assembles as a dimer in the plasma membrane. Addition of Ca(2+) decreases the distance between the cytoplasmic loops of NCX pairs. The Ca(2+)-dependent movements detected between paired NCXs were abolished by mutating the Ca(2+) coordination sites in CBD1 (D421A, E451A, and D500V), whereas disruption of the primary Ca(2+) coordination site in CBD2 (E516L) had no effect. Thus, the Ca(2+)-induced conformational changes of NCX dimers arise from the movement of CBD1. FRET studies of CBD1, CBD2, and CBD1-CBD2 peptides displayed Ca(2+)-dependent movements with different apparent affinities. CBD1-CBD2 showed a Ca(2+)-dependent phenotype mirroring full-length NCX but distinct from both CBD1 and CBD2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Philipson KD, et al. The Na+/Ca2+ exchange molecule: An overview. Ann N Y Acad Sci. 2002;976:1–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
