

The aspartate-semialdehyde dehydrogenase of Edwardsiella ictaluri and its use as balanced-lethal system in fish vaccinology
- PMID: 21209920
- PMCID: PMC3012122
- DOI: 10.1371/journal.pone.0015944
The aspartate-semialdehyde dehydrogenase of Edwardsiella ictaluri and its use as balanced-lethal system in fish vaccinology
Abstract
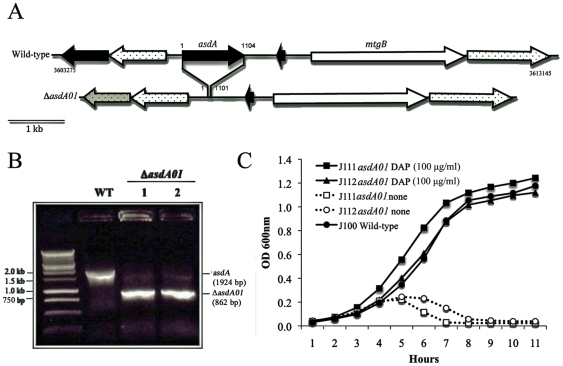
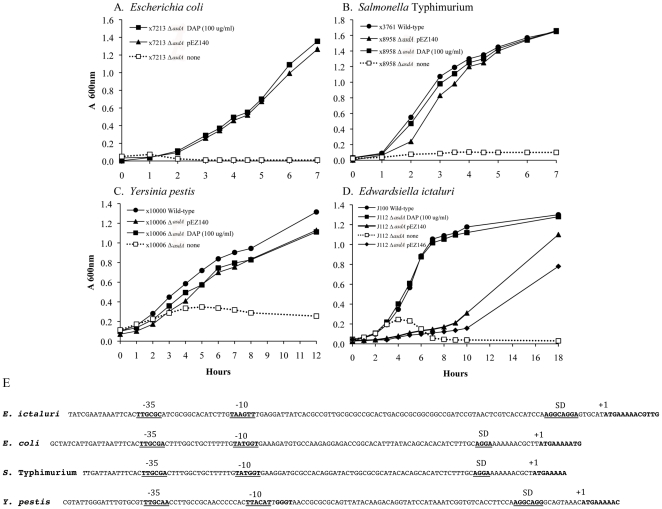
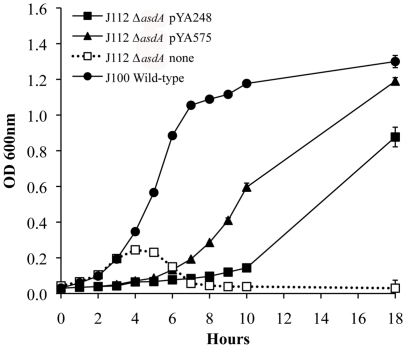
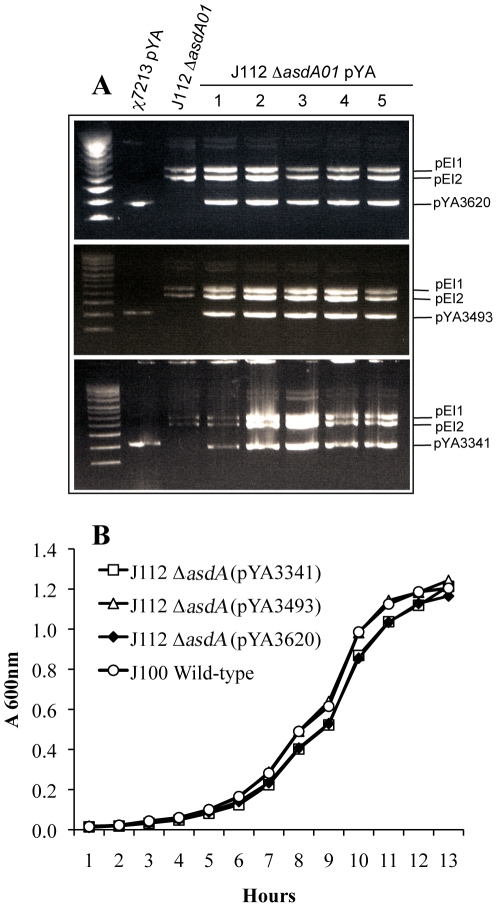
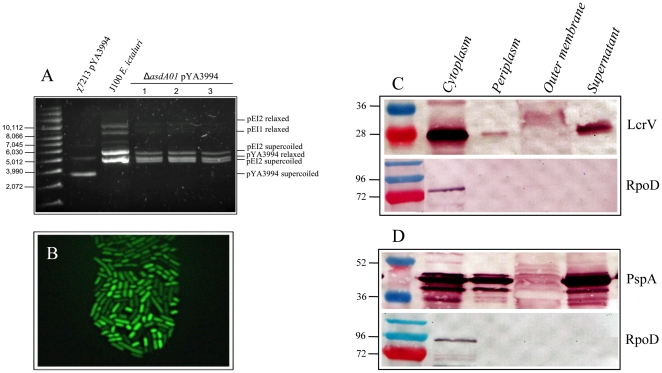
asdA mutants of gram-negative bacteria have an obligate requirement for diaminopimelic acid (DAP), which is an essential constituent of the peptidoglycan layer of the cell wall of these organisms. In environments deprived of DAP, i.e., animal tissues, they will undergo lysis. Deletion of the asdA gene has previously been exploited to develop antibiotic-sensitive strains of live attenuated recombinant bacterial vaccines. Introduction of an Asd(+) plasmid into a ΔasdA mutant makes the bacterial strain plasmid-dependent. This dependence on the Asd(+) plasmid vector creates a balanced-lethal complementation between the bacterial strain and the recombinant plasmid. E. ictaluri is an enteric gram-negative fish pathogen that causes enteric septicemia in catfish. Because E. ictaluri is a nasal/oral invasive intracellular pathogen, this bacterium is a candidate to develop a bath/oral live recombinant attenuated Edwardsiella vaccine (RAEV) for the catfish aquaculture industry. As a first step to develop an antibiotic-sensitive RAEV strain, we characterized and deleted the E. ictaluri asdA gene. E. ictaluri ΔasdA01 mutants exhibit an absolute requirement for DAP to grow. The asdA gene of E. ictaluri was complemented by the asdA gene from Salmonella. Several Asd(+) expression vectors with different origins of replication were transformed into E. ictaluri ΔasdA01. Asd(+) vectors were compatible with the pEI1 and pEI2 E. ictaluri native plasmids. The balanced-lethal system was satisfactorily evaluated in vivo. Recombinant GFP, PspA, and LcrV proteins were synthesized by E. ictaluri ΔasdA01 harboring Asd(+) plasmids. Here we constructed a balanced-lethal system, which is the first step to develop an antibiotic-sensitive RAEV for the aquaculture industry.
Conflict of interest statement
Figures

Similar articles
-
Construction and Evaluation of Recombinant Attenuated Edwardsiella piscicida Vaccine (RAEV) Vector System Encoding Ichthyophthirius multifiliis (Ich) Antigen IAG52B.Front Immunol. 2022 Jan 25;12:802760. doi: 10.3389/fimmu.2021.802760. eCollection 2021. Front Immunol. 2022. PMID: 35145512 Free PMC article.
-
Phenotype, virulence and immunogenicity of Edwardsiella ictaluri cyclic adenosine 3',5'-monophosphate receptor protein (Crp) mutants in catfish host.Fish Shellfish Immunol. 2011 Dec;31(6):1142-53. doi: 10.1016/j.fsi.2011.10.009. Epub 2011 Oct 14. Fish Shellfish Immunol. 2011. PMID: 22015784
-
Asd-based balanced-lethal system in attenuated Edwardsiella tarda to express a heterologous antigen for a multivalent bacterial vaccine.Fish Shellfish Immunol. 2013 May;34(5):1188-94. doi: 10.1016/j.fsi.2013.01.027. Epub 2013 Feb 27. Fish Shellfish Immunol. 2013. PMID: 23454428
-
Roles of plasmid-encoded proteins, EseH, EseI and EscD in invasion, replication and virulence of Edwardsiella ictaluri.Vet Microbiol. 2013 Sep 27;166(1-2):233-41. doi: 10.1016/j.vetmic.2013.05.023. Epub 2013 Jun 14. Vet Microbiol. 2013. PMID: 23850444
-
Molecular genetic analysis of the region containing the essential Pseudomonas aeruginosa asd gene encoding aspartate-beta-semialdehyde dehydrogenase.Microbiology (Reading). 1997 Mar;143 ( Pt 3):899-907. doi: 10.1099/00221287-143-3-899. Microbiology (Reading). 1997. PMID: 9084174
Cited by
-
Fur-regulated iron uptake system of Edwardsiella ictaluri and its influence on pathogenesis and immunogenicity in the catfish host.Infect Immun. 2012 Aug;80(8):2689-703. doi: 10.1128/IAI.00013-12. Epub 2012 May 21. Infect Immun. 2012. PMID: 22615248 Free PMC article.
-
C500 variants conveying complete mucosal immunity against fatal infections of pigs with Salmonella enterica serovar Choleraesuis C78-1 or F18+ Shiga toxin-producing Escherichia coli.Front Microbiol. 2023 Sep 14;14:1210358. doi: 10.3389/fmicb.2023.1210358. eCollection 2023. Front Microbiol. 2023. PMID: 37779705 Free PMC article.
-
Inflammatory effects of Edwardsiella ictaluri lipopolysaccharide modifications in catfish gut.Infect Immun. 2014 Aug;82(8):3394-404. doi: 10.1128/IAI.01697-14. Epub 2014 May 27. Infect Immun. 2014. PMID: 24866806 Free PMC article.
-
Generation of influenza virus from avian cells infected by Salmonella carrying the viral genome.PLoS One. 2015 Mar 5;10(3):e0119041. doi: 10.1371/journal.pone.0119041. eCollection 2015. PLoS One. 2015. PMID: 25742162 Free PMC article.
-
The Asd(+)-DadB(+) dual-plasmid system offers a novel means to deliver multiple protective antigens by a recombinant attenuated Salmonella vaccine.Infect Immun. 2012 Oct;80(10):3621-33. doi: 10.1128/IAI.00620-12. Epub 2012 Aug 6. Infect Immun. 2012. PMID: 22868499 Free PMC article.
References
-
- Viola RE. The central enzymes of the aspartate family of amino acid biosynthesis. Acc Chem Res. 2001;34:339–349. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
