

Review
doi: 10.1016/j.cell.2010.11.056.
Epigenetics in alternative pre-mRNA splicing
Affiliations
- PMID: 21215366
- PMCID: PMC3038581
- DOI: 10.1016/j.cell.2010.11.056
Item in Clipboard
Review
Epigenetics in alternative pre-mRNA splicing
Cell.
.
Abstract
Alternative splicing plays critical roles in differentiation, development, and disease and is a major source for protein diversity in higher eukaryotes. Analysis of alternative splicing regulation has traditionally focused on RNA sequence elements and their associated splicing factors, but recent provocative studies point to a key function of chromatin structure and histone modifications in alternative splicing regulation. These insights suggest that epigenetic regulation determines not only what parts of the genome are expressed but also how they are spliced.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
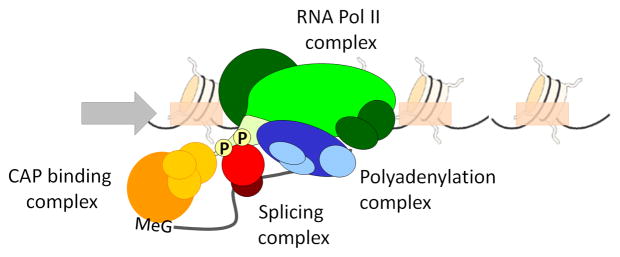
RNA polymerase II (green) recruits RNA processing factors such as the 5′ cap-binding complex (CAP) (yellow), splicing and pre-spliceosome factors (red) and the polyadenylation complex (blue) in the context of nucleosome-containing chromatin. Recruitment of RNA processing factors occurs via the RNA Pol II C-terminal domain (CTD; light green) and much of RNA processing occurs co-transcriptionally.
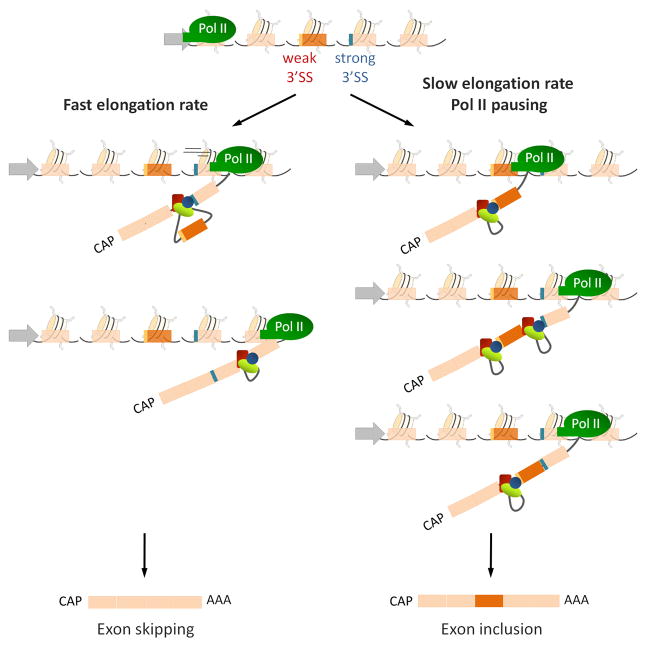
Rapid elongation of RNA Polymerase II (Pol II) leads to simultaneous availability to the splicing machinery of a weak (red) and a strong (blue) splice site which compete for the recruitment of splicing factors (red, blue and green ovals) resulting in skipping of the weaker exon (orange rectangle). Pausing or slowing down of the RNA Pol II favors the recruitment of the splicing machinery to the first transcribed, weaker exon leading to its subsequent inclusion in a “first served, first committed” model.
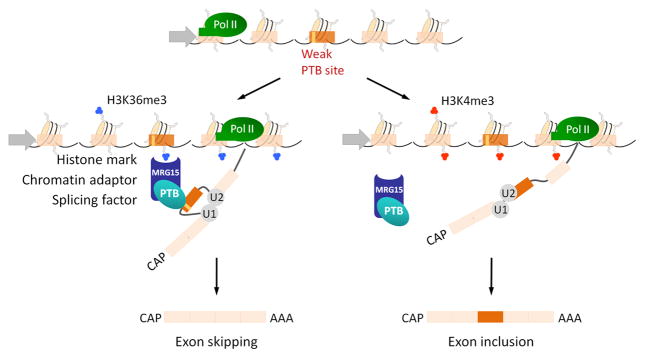
Histone modifications along the gene determine the binding of an adaptor protein which reads specific histone marks and in turn recruits splicing factors. In the case of exons whose alternative splicing is dependent on polypyrimidine tract binding protein (PTB) splicing factor, high levels of trimethylated histone 3 lysine 36 (H3K36me3, blue) attract the chromatin-binding factor MRG15 that acts as an adaptor protein and by protein-protein interaction helps to recruit PTB to its weaker binding site inducing exon skipping. If the PTB-dependent gene is hypermethylated in H3K4me3 (red), MRG15 does not accumulate along the gene, and PTB is not recruited to its target pre-mRNA, thus favoring exon inclusion.
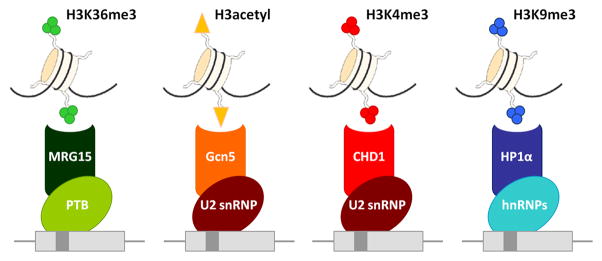
Several histone modification-binding chromatin proteins interact with splicing factors (Luco et al., 2010; Sims et al., 2007; Gunderson and Johnson, 2009; Piacentini et al., 2009; Loomis et al., 2009).
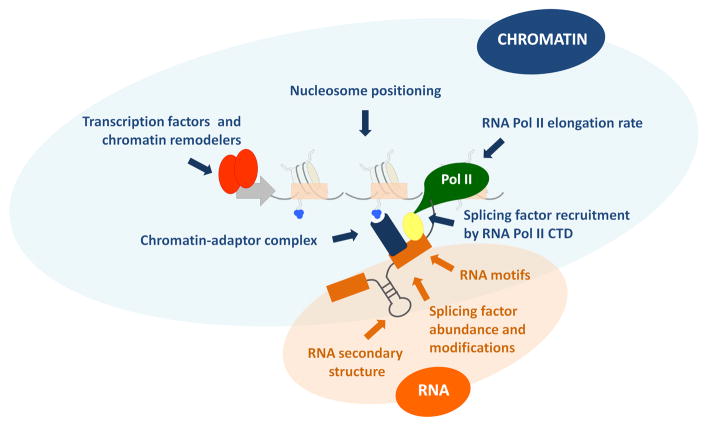
Alternative splicing patterns are determined by a combination of parameters including cis-acting RNA regulatory elements and RNA secondary structures (highlighted in orange) together with transcriptional and chromatin properties (highlighted in blue) that modulate the recruitment of splicing factors to the pre-mRNA.
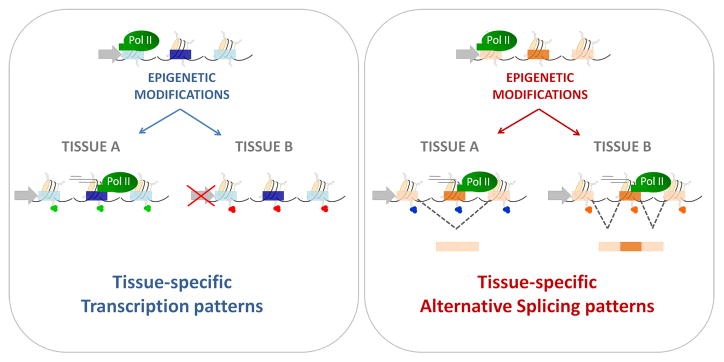
The combination of histone modifications along a gene establishes and maintains tissue-specific transcription patterns (left panel), as well as heritable tissue-specific alternative splicing patterns (right panel).
References
-
- Aebi M, Weissmann C. Precision and orderliness in splicing. Trends Genet. 1987;3:102–107.
-
- Allo M, Buggiano V, Fededa JP, Petrillo E, Schor I, de la Mata M, Agirre E, Plass M, Eyras E, Abou Elela S, Klinck R, Chabot B, Kornblihtt AR. Control of alternative splicing through siRNA-mediated transcriptional gene silencing. Nat Struct Mol Biol. 2009;16:717–724. - PubMed
-
- Attanasio C, David A, Neerman-Arbez M. Outcome of donor splice site mutations accounting for congenital afibrinogenemia reflects order of intron removal in the fibrinogen alpha gene (FGA) Blood. 2003;101:1851–1856. - PubMed
-
- Auboeuf D, Honig A, Berget SM, O’Malley BW. Coordinate regulation of transcription and splicing by steroid receptor coregulators. Science. 2002;298:416–419. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
