

Review
doi: 10.1128/AEM.02473-10.
Epub 2011 Jan 7.
Ecological aspects of the distribution of different autotrophic CO2 fixation pathways
Affiliations
- PMID: 21216907
- PMCID: PMC3067309
- DOI: 10.1128/AEM.02473-10
Item in Clipboard
Review
Ecological aspects of the distribution of different autotrophic CO2 fixation pathways
Appl Environ Microbiol.
2011 Mar.
Abstract
Autotrophic CO(2) fixation represents the most important biosynthetic process in biology. Besides the well-known Calvin-Benson cycle, five other totally different autotrophic mechanisms are known today. This minireview discusses the factors determining their distribution. As will be made clear, the observed diversity reflects the variety of the organisms and the ecological niches existing in nature.
Figures
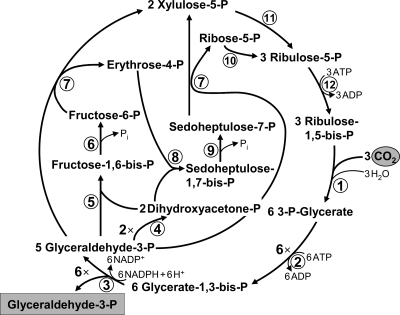
The reductive pentose phosphate (Calvin-Benson) cycle. Enzymes: 1, ribulose-1,5-bisphosphate carboxylase/oxygenase; 2, 3-phosphoglycerate kinase; 3, glyceraldehyde-3-phosphate dehydrogenase; 4, triose-phosphate isomerase; 5, fructose-bisphosphate aldolase; 6, fructose-bisphosphate phosphatase; 7, transketolase; 8, sedoheptulose-bisphosphate aldolase; 9, sedoheptulose-bisphosphate phosphatase; 10, ribose-phosphate isomerase; 11, ribulose-phosphate epimerase; and 12, phosphoribulokinase.
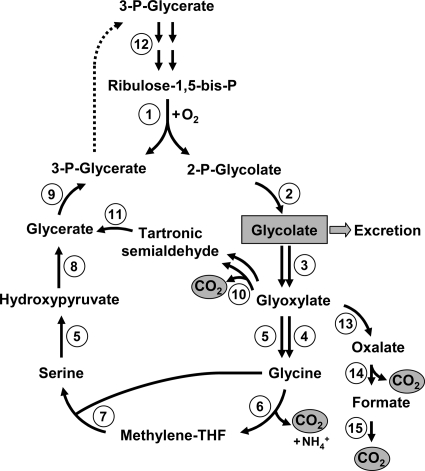
Photorespiration. RubisCO-catalyzed oxygenation of ribulose-1,5-bisphosphate (enzyme 1) leads to the formation of 3-phosphoglycerate and 2-phosphoglycolate. 3-Phosphoglycerate returns to the CB cycle, whereas 2-phosphoglycolate is converted to glycolate by 2-phosphoglycolate phosphatase (enzyme 2). Glycolate can either be excreted or oxidized to glyoxylate by glycolate dehydrogenase or glycolate oxidase (enzyme 3). In the classical pathway functioning in, e.g., plants, glyoxylate is converted to glycine by glutamate-glyoxylate aminotransferase (enzyme 4) and serine-glyoxylate transaminase (enzyme 5). One molecule of glycine is then oxidized by the glycine decarboxylase complex (enzymes 6) to methylene-tetrahydrofolate (THF), which reacts with the second glycine molecule in the reaction, catalyzed by serine hydroxymethyltransferase (enzyme 7). The resulting serine is transaminated to hydroxypyruvate (enzyme 5) and then converted to 3-phosphoglycerate by hydroxypyruvate reductase (enzyme 8) and glycerate kinase (enzyme 9). An alternative pathway of glyoxylate assimilation is the condensation of two glyoxylate molecules giving rise to tartronic semialdehyde (glyoxylate carboligase [enzyme 10]). Tartronic semialdehyde reductase reaction (enzyme 11) leads to glycerate, and the latter is converted to 3-phosphoglycerate. Ribulose-1,5-bisphosphate regeneration proceeds via the CB cycle reactions (step 12). In some cases, glyoxylate can also be completely oxidized to CO2 in reactions catalyzed by hydroxyacid dehydrogenase (enzyme 13), oxalate decarboxylase (enzyme 14), and formate dehydrogenase (enzyme 15). All of these pathways can be found in cyanobacteria (27).
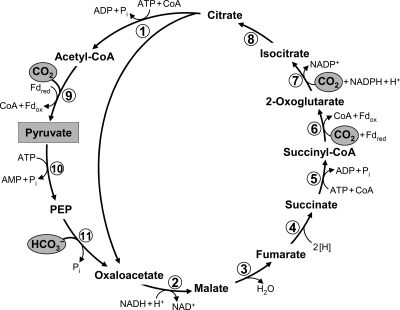
The reductive citric acid (Arnon-Buchanan) cycle as it functions in green sulfur bacteria (33, 52). The pathway of acetyl-CoA assimilation to pyruvate, phosphoenolpyruvate (PEP), and oxaloacetate is shown as well. For deviations from this variant of the cycle, see the text. Enzymes: 1, ATP-citrate lyase; 2, malate dehydrogenase; 3, fumarate hydratase; 4, fumarate reductase (natural electron donor is not known); 5, succinyl-CoA synthetase; 6, ferredoxin (Fd)-dependent 2-oxoglutarate synthase; 7, isocitrate dehydrogenase; 8, aconitate hydratase; 9, Fd-dependent pyruvate synthase; 10, PEP synthase; 11, PEP carboxylase.
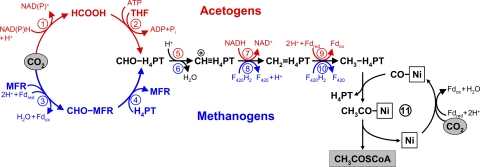
The reductive acetyl-CoA (Wood-Ljungdahl) pathway. One CO2 molecule is reduced to carbon monoxide bound to a nickel atom in the active center of CO dehydrogenase, and another is reduced to a methyl group bound to the carrier tetrahydropterin; subsequent methyl transfer to nickel-bound CO leads to acetyl-CoA synthesis. The upper part (red) shows the variant of the pathway functioning in acetogens, and the lower part (blue) depicts the pathway in methanogens. Fd, ferredoxin; THF, tetrahydrofolate; H4TPT, tetrahydropterin (THF in acetogens and tetrahydromethanopterin or tetrahydrosarcinopterin in methanogens); MFR, methanofuran; F420, coenzyme F420. Enzymes: 1, formate dehydrogenase; 2; formyl-THF synthetase; 3, formyl-MFR dehydrogenase; 4, formyl-MFR:tetrahydromethanopterin formyltransferase; 5, methenyl-THF cyclohydrolase; 6, methenyl-tetrahydromethanopterin cyclohydrolase; 7, methylene-THF dehydrogenase; 8, methylene-tetrahydromethanopterin dehydrogenase; 9, methylene-THF reductase; 10, methylene-tetrahydromethanopterin reductase; 11, CO dehydrogenase/acetyl-CoA synthase.
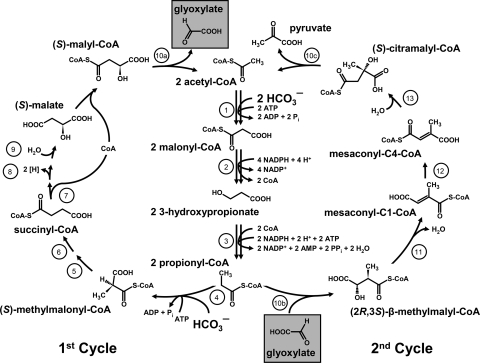
The 3-hydroxypropionate (Fuchs-Holo) bi-cycle (130). Enzymes: 1, acetyl-CoA carboxylase; 2, malonyl-CoA reductase; 3, propionyl-CoA synthase; 4, propionyl-CoA carboxylase; 5, methylmalonyl-CoA epimerase; 6, methylmalonyl-CoA mutase; 7, succinyl-CoA:(S)-malate-CoA transferase; 8, succinate dehydrogenase; 9, fumarate hydratase; 10a, -b, and -c, trifunctional (S)-malyl-CoA (a)/β-methylmalyl-CoA (b)/(S)-citramalyl-CoA lyase (c); 11, mesaconyl-C1-CoA hydratase; 12, mesaconyl-CoA C1-C4 CoA transferase; 13, mesaconyl-C4-CoA hydratase. (Reprinted from reference with permission.)
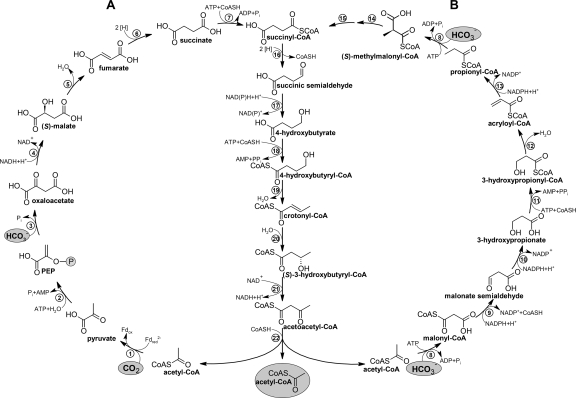
The 4-hydroxybutyrate cycles of autotrophic CO2 fixation (14). (A) The dicarboxylate/4-hydroxybutyrate cycle functioning in Desulfurococcales and Thermoproteales; (B) the 3-hydroxypropionate/4-hydroxybutyrate cycle functioning in Sulfolobales. Note that succinyl-CoA reductase in Thermoproteales and Sulfolobales uses NADPH (13, 62, 81, 82) and probably reduced ferredoxin (Fdred) in Desulfurococcales (15, 46). Enzymes: 1, pyruvate synthase; 2, pyruvate:water dikinase; 3, PEP carboxylase; 4, malate dehydrogenase; 5, fumarate hydratase; 6, fumarate reductase (natural electron acceptor is not known); 7, succinyl-CoA synthetase; 8, acetyl-CoA/propionyl-CoA carboxylase; 9, malonyl-CoA reductase; 10, malonic semialdehyde reductase; 11, 3-hydroxypropionate-CoA ligase; 12, 3-hydroxypropionyl-CoA dehydratase; 13, acryloyl- CoA reductase; 14, methylmalonyl-CoA epimerase; 15, methylmalonyl-CoA mutase; 16, succinyl-CoA reductase; 17, succinic semialdehyde reductase; 18, 4-hydroxybutyrate-CoA ligase; 19, 4-hydroxybutyryl-CoA dehydratase; 20, crotonyl-CoA hydratase; 21, (S)-3-hydroxybutyryl-CoA dehydrogenase (NAD+); 22, acetoacetyl-CoA β-ketothiolase. Fd, ferredoxin. (Adapted from reference .)
References
-
- Anderson, L. E. 1971. Chloroplast and cytoplasmic enzymes. II. Pea leaf triose phosphate isomerases. Biochim. Biophys. Acta 235:237-244. - PubMed
-
- Aoshima, M. 2007. Novel enzyme reactions related to the citric acid cycle: phylogenetic/functional implications and biotechnological applications. Appl. Microbiol. Biotechnol. 75:249-255. - PubMed
-
- Aoshima, M., M. Ishii, and Y. Igarashi. 2004. A novel enzyme, citryl-CoA synthetase, catalysing the first step of the citrate cleavage reaction in Hydrogenobacter thermophilus TK-6. Mol. Microbiol. 52:751-761. - PubMed
-
- Aoshima, M., M. Ishii, and Y. Igarashi. 2004. A novel enzyme, citryl-CoA lyase, catalysing the second step of the citrate cleavage reaction in Hydrogenobacter thermophilus TK-6. Mol. Microbiol. 52:763-770. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
