

Thermodynamics and mechanics of membrane curvature generation and sensing by proteins and lipids
- PMID: 21219150
- PMCID: PMC4205088
- DOI: 10.1146/annurev.physchem.012809.103450
Thermodynamics and mechanics of membrane curvature generation and sensing by proteins and lipids
Abstract
Research investigating lipid membrane curvature generation and sensing is a rapidly developing frontier in membrane physical chemistry and biophysics. The fast recent progress is based on the discovery of a plethora of proteins involved in coupling membrane shape to cellular membrane function, the design of new quantitative experimental techniques to study aspects of membrane curvature, and the development of analytical theories and simulation techniques that allow a mechanistic interpretation of quantitative measurements. The present review first provides an overview of important classes of membrane proteins for which function is coupled to membrane curvature. We then survey several mechanisms that are assumed to underlie membrane curvature sensing and generation. Finally, we discuss relatively simple thermodynamic/mechanical models that allow quantitative interpretation of experimental observations.
Figures
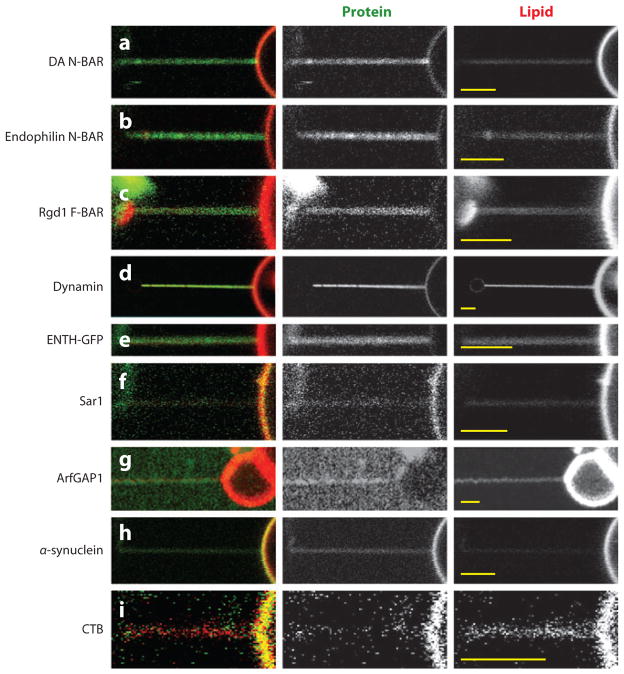
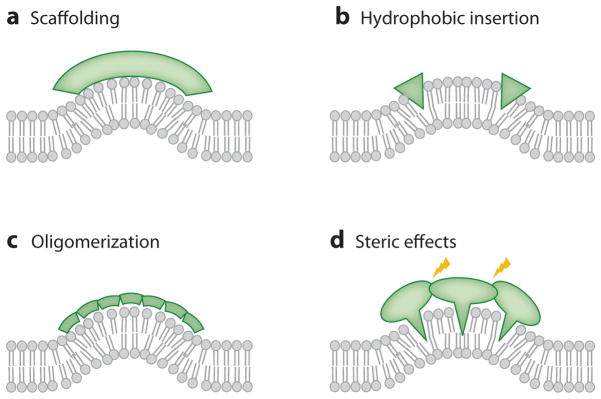
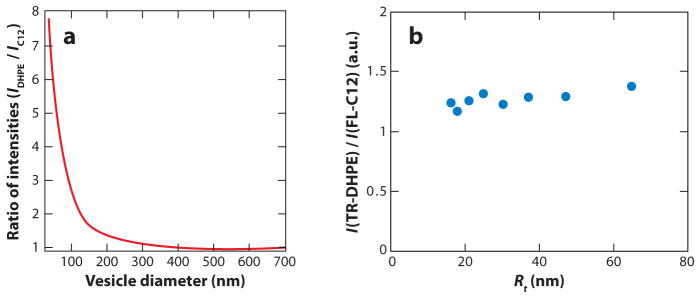
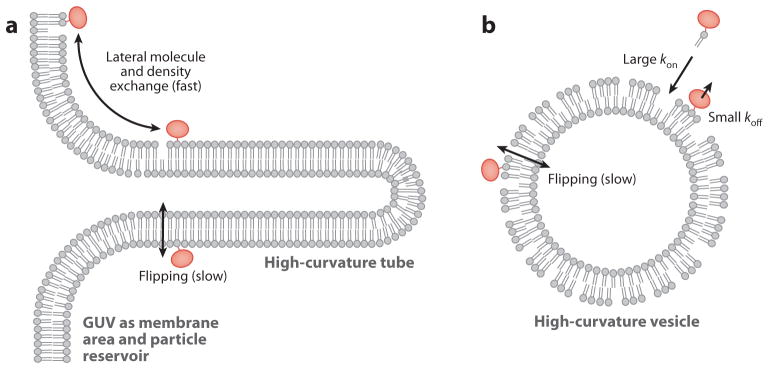
References
-
- Zimmerberg J, Kozlov MM. How proteins produce cellular membrane curvature. Nat Rev Mol Cell Biol. 2006;7:9–19. - PubMed
-
- McMahon HT, Gallop JL. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature. 2005;438:590–96. - PubMed
-
- Sweitzer SM, Hinshaw JE. Dynamin undergoes a GTP-dependent conformational change causing vesiculation. Cell. 1998;93:1021–29. - PubMed
-
- Takei K, Slepnev VI, Haucke V, De Camilli P. Functional partnership between amphiphysin and dynamin in clathrin-mediated endocytosis. Nat Cell Biol. 1999;1:33–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
