

A mouse model of the fragile X premutation: effects on behavior, dendrite morphology, and regional rates of cerebral protein synthesis
- PMID: 21220020
- PMCID: PMC3150744
- DOI: 10.1016/j.nbd.2011.01.008
A mouse model of the fragile X premutation: effects on behavior, dendrite morphology, and regional rates of cerebral protein synthesis
Abstract
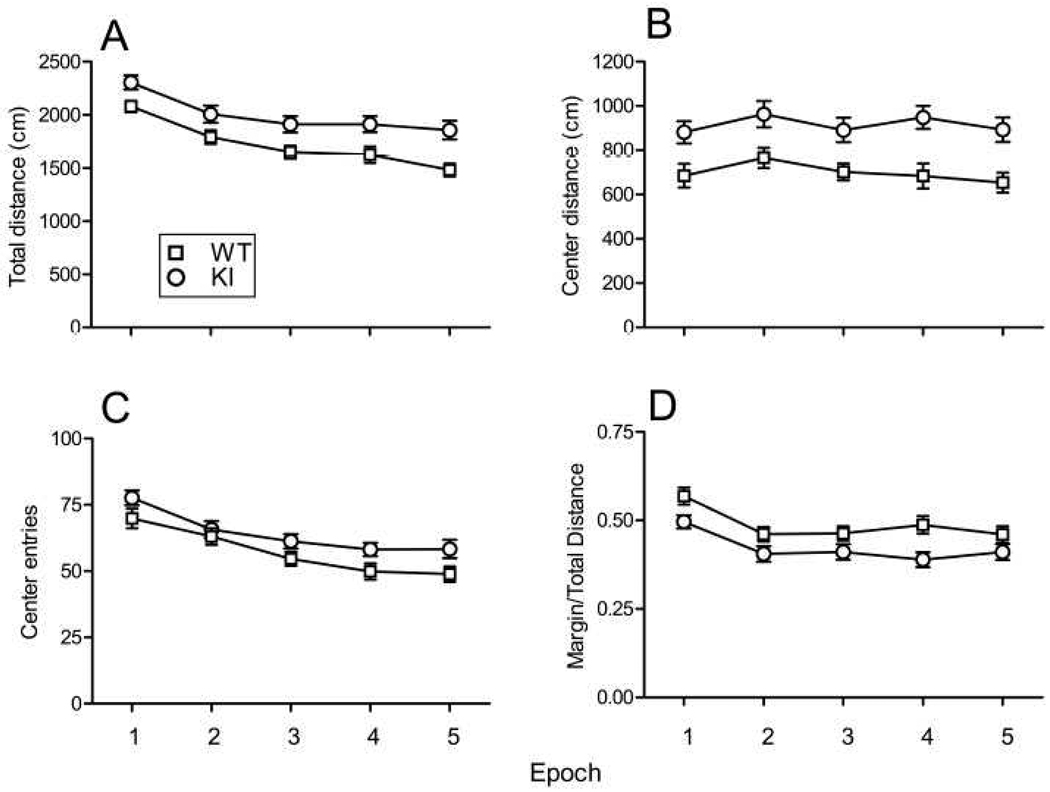
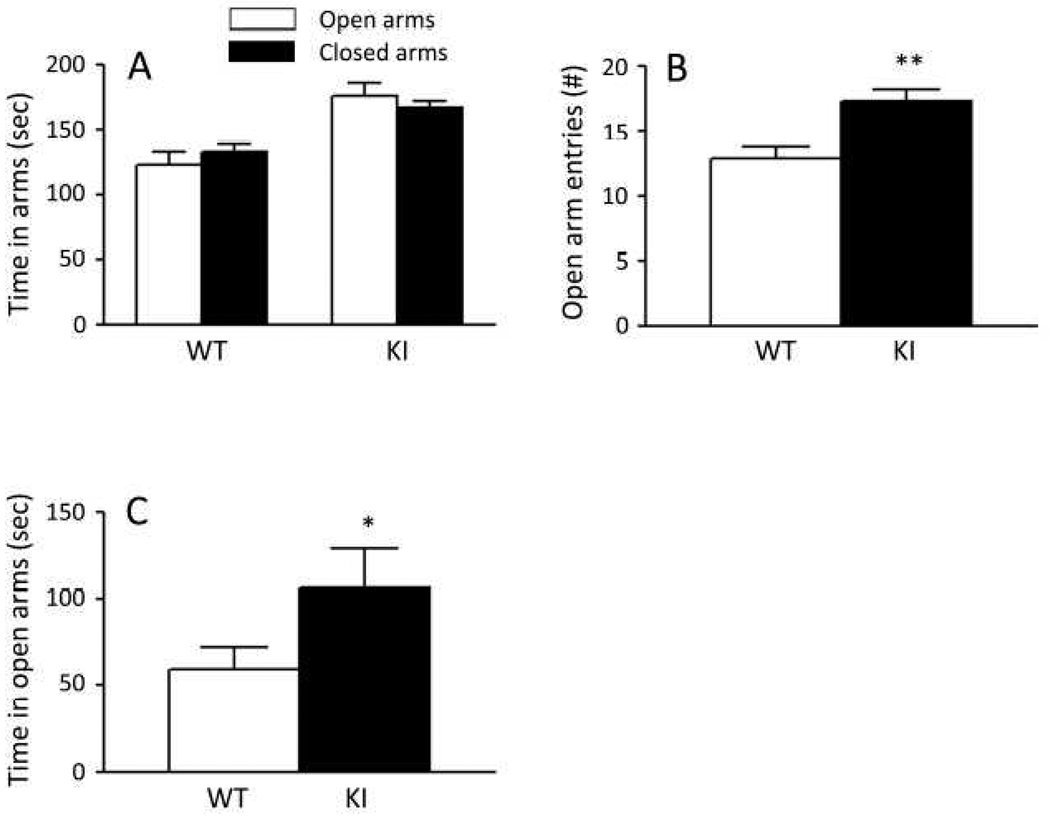
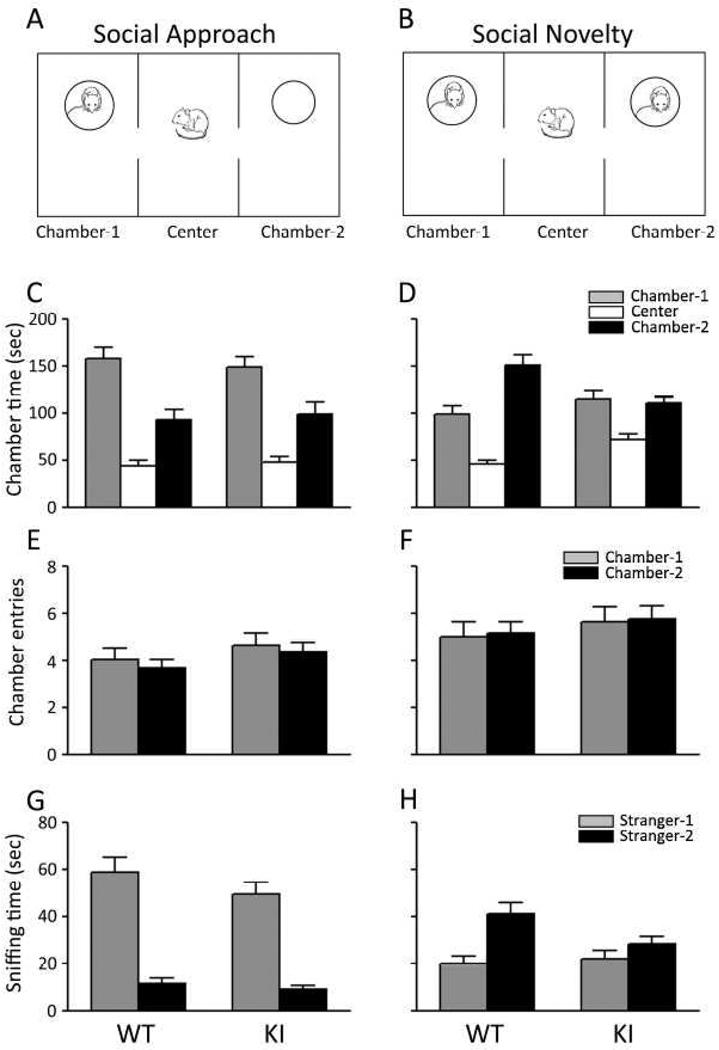
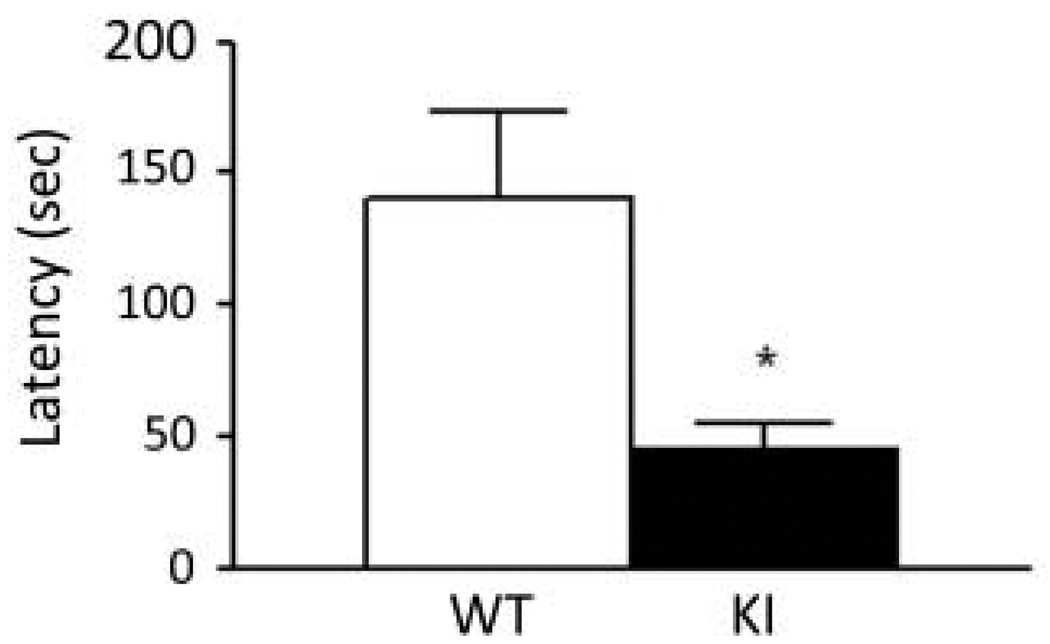
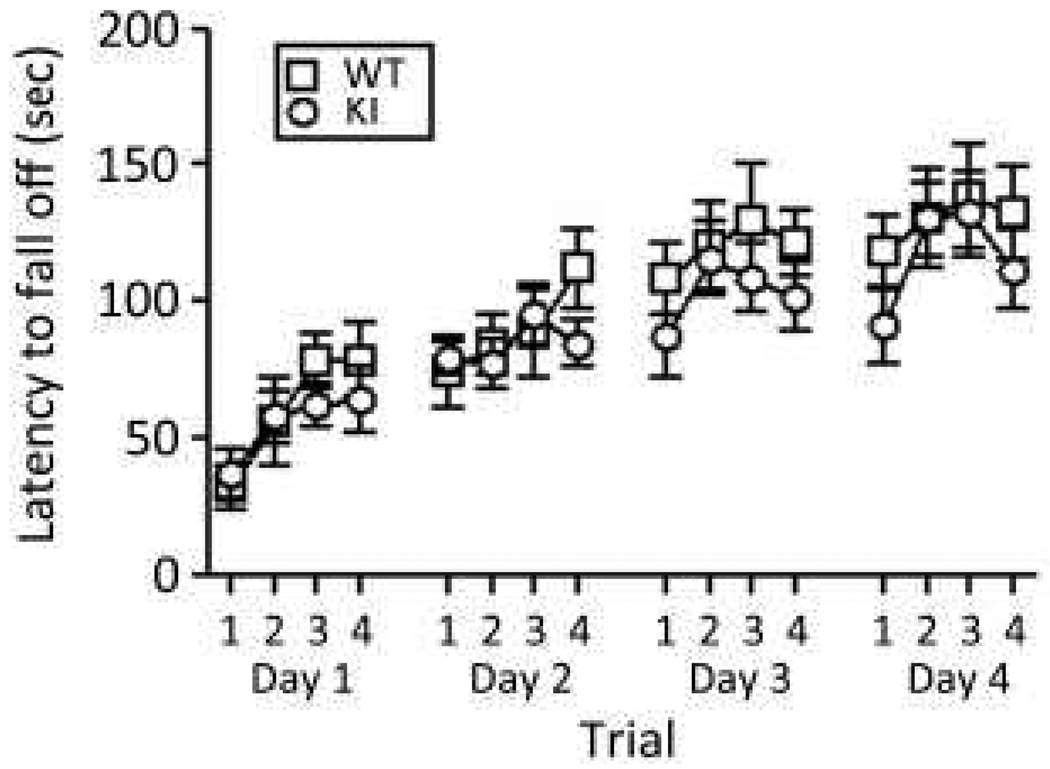
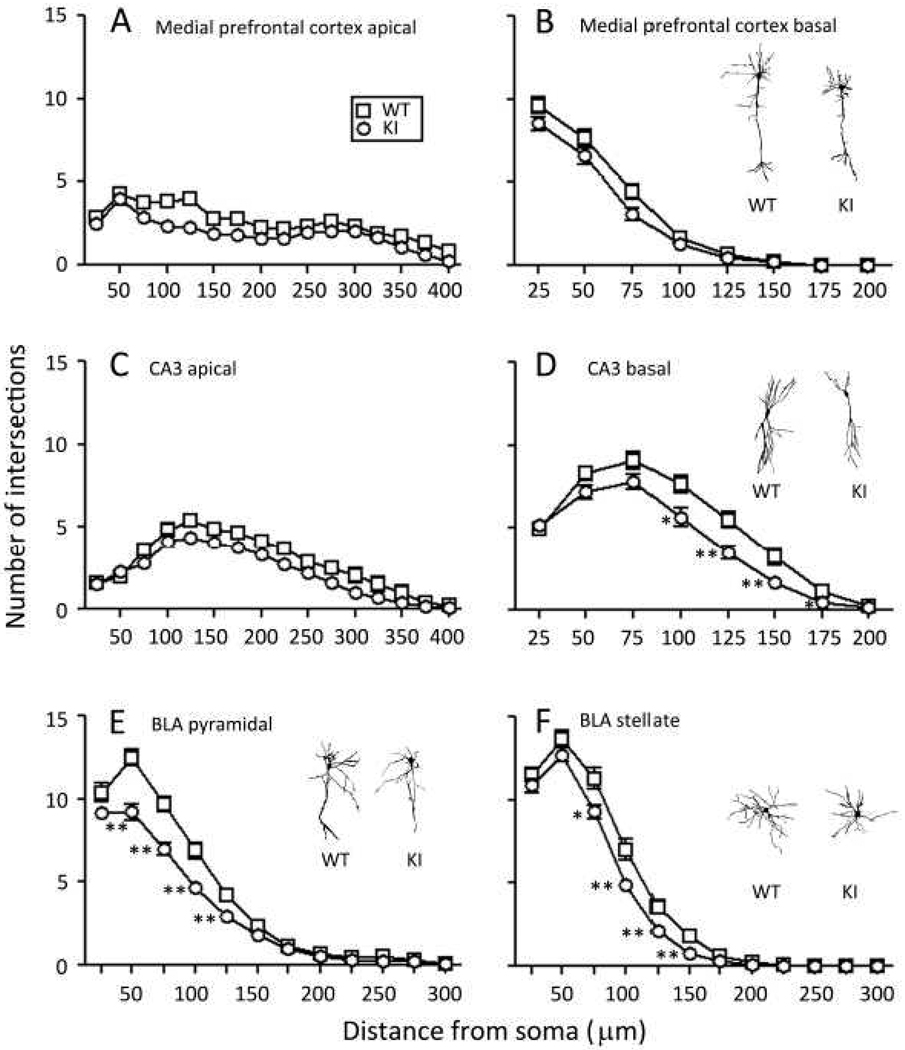
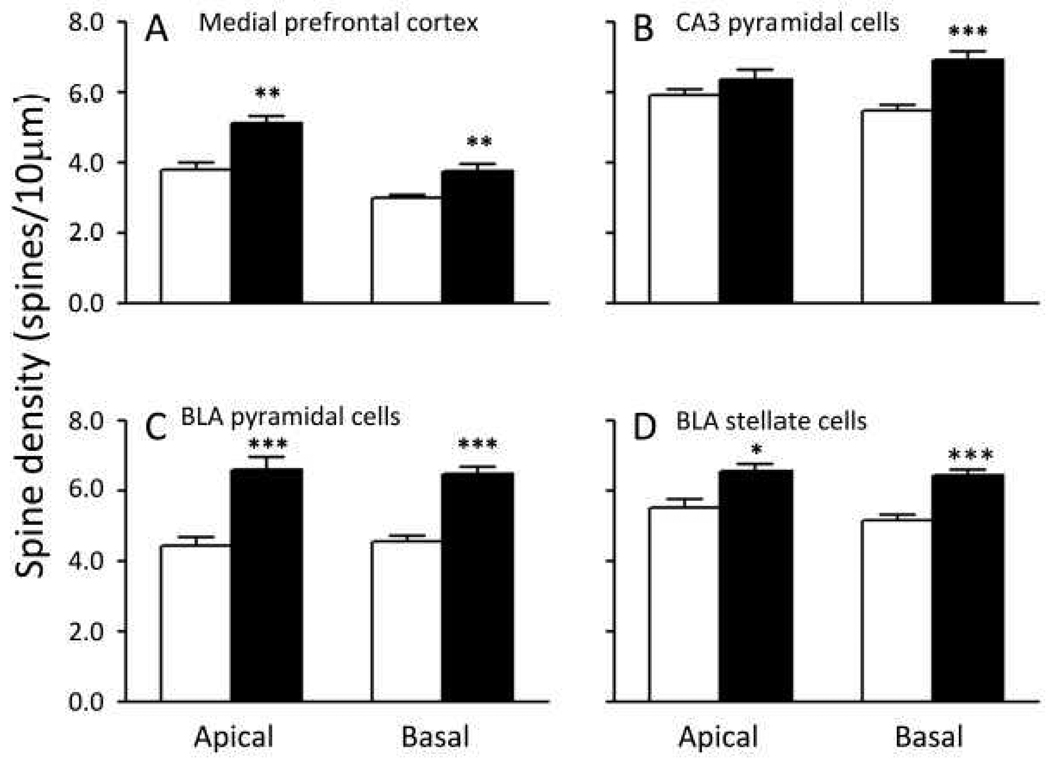
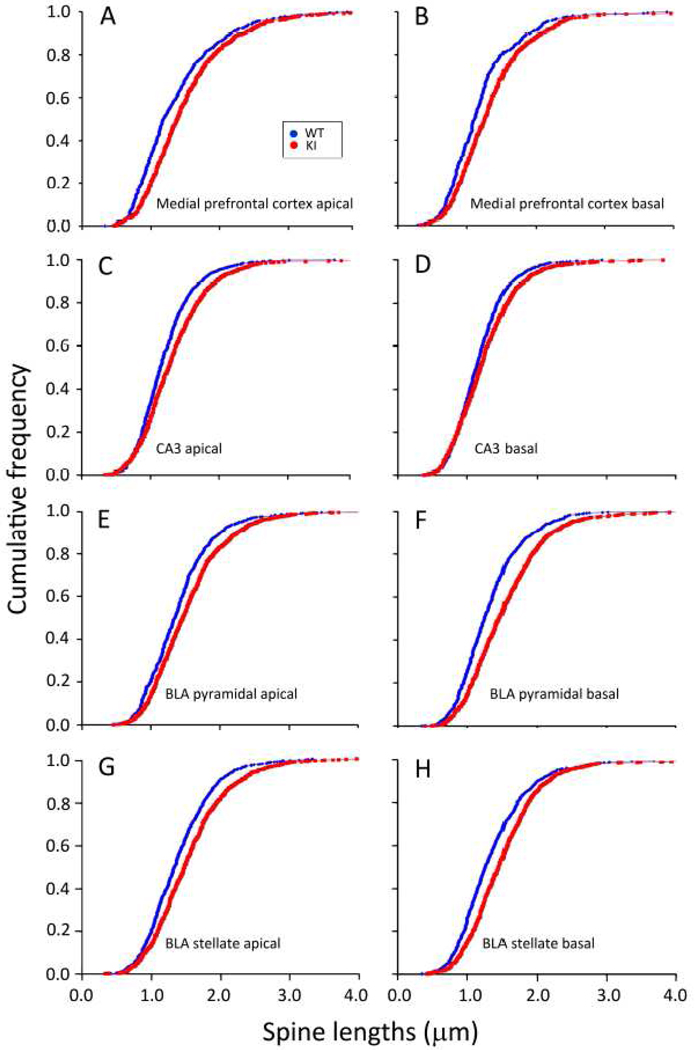
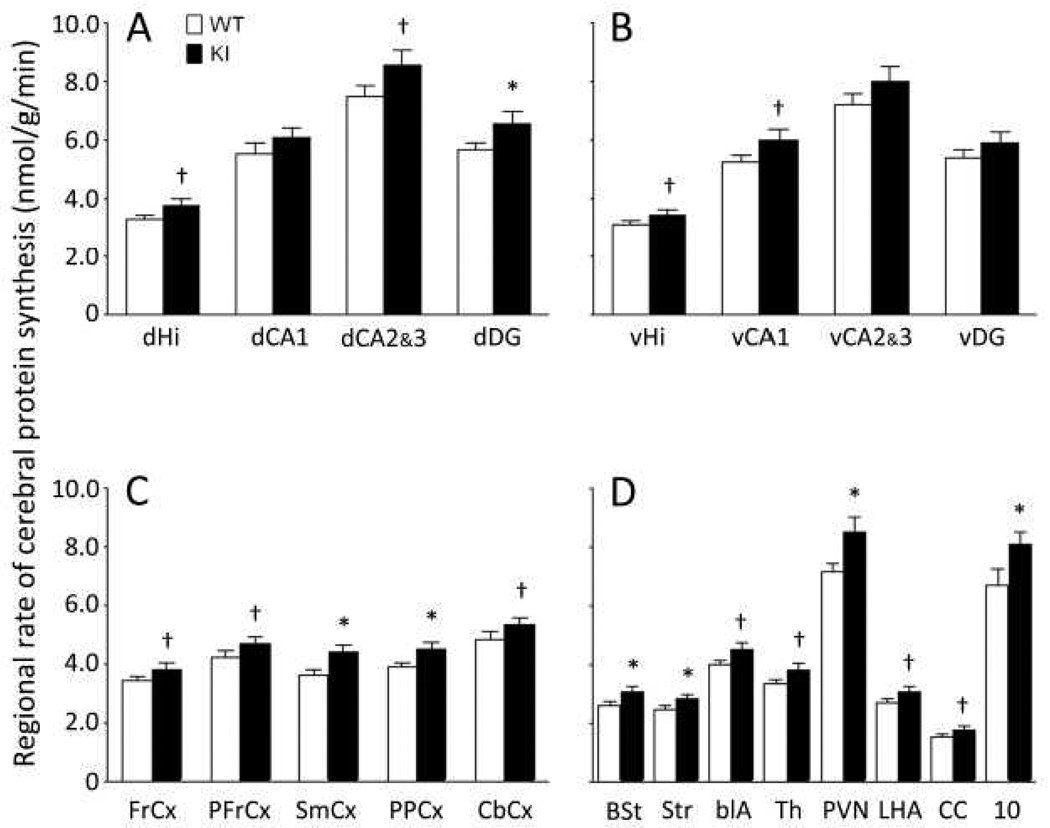
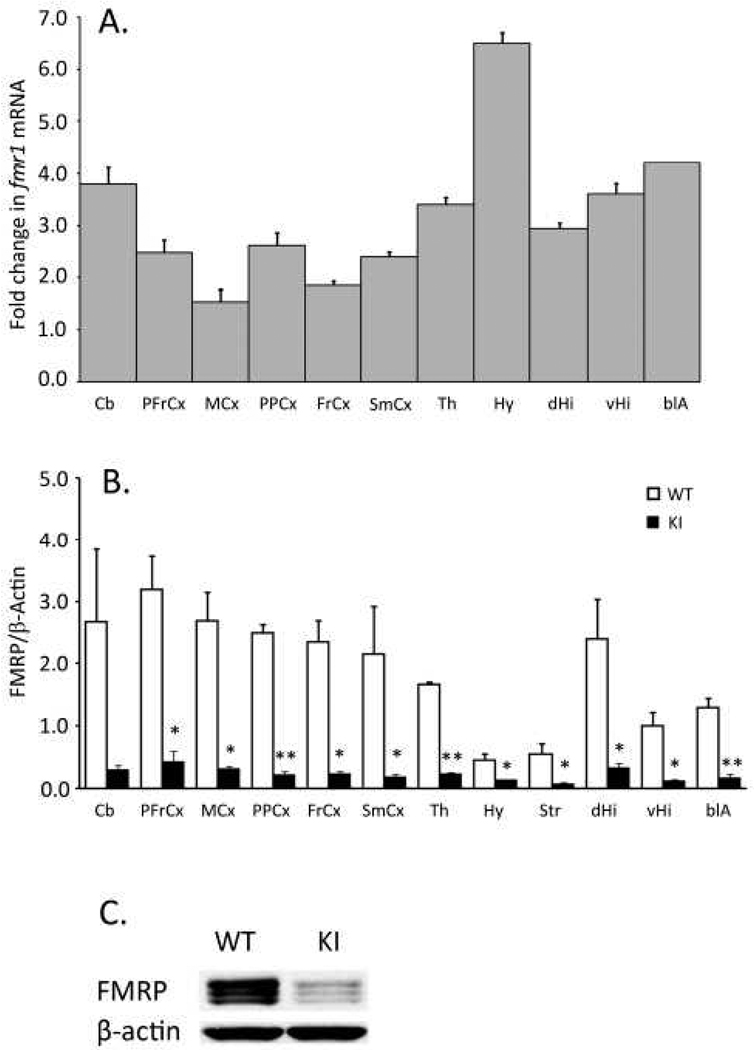
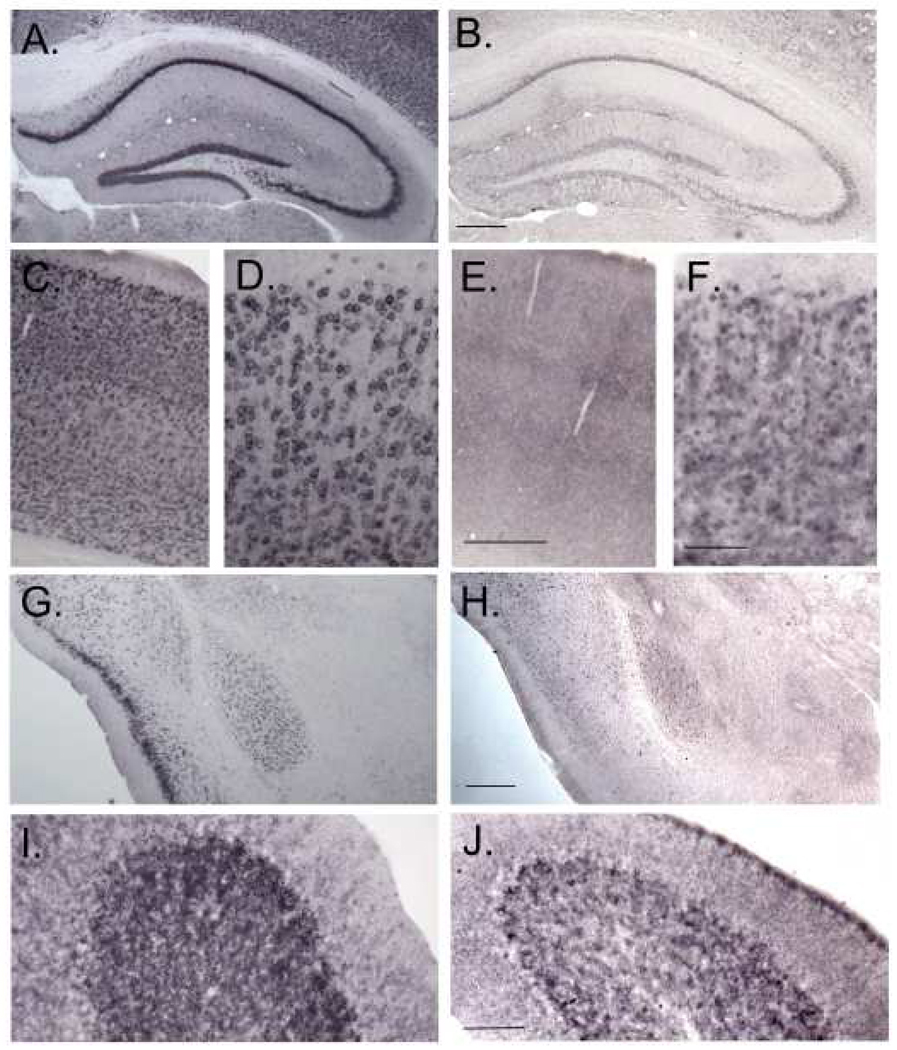
Carriers of FMR1 premutation alleles have 55-200 CGG repeats in the 5' untranslated region of the gene. These individuals are at risk for fragile X associated primary ovarian insufficiency (females) and, in late life, fragile X associated tremor and ataxia syndrome (males, and to a lesser extent, females). Premutation carrier status can also be associated with autism spectrum disorder, attention deficit hyperactivity disorder, and some cognitive deficits. In premutation carriers, FMR1 mRNA levels are often higher than those with normal sized alleles. In contrast, in subjects with full mutation alleles, (>200 repeats) the FMR1 gene is silenced and FMR1 mRNA and its product, FMRP, are absent. We have studied a male knock-in (KI) mouse model of the fragile X premutation (120-140 repeats) during young adulthood. In comparison to wild type, KI mice were hyperactive, exhibited less anxiety in both the open field and the elevated zero maze, were impaired on the passive avoidance test, and showed some subtle deficits on a test of social interaction. Motor learning as assessed by the rotarod test was normal. Dendritic arbors were less complex and spine densities and lengths increased in medial prefrontal cortex, basal lateral amygdala, and hippocampus compared with wild type. Regional rates of cerebral protein synthesis measured in vivo in KI mice were increased. KI mice also had elevated levels of Fmr1 mRNA and decreased levels of FMRP. Our results highlight similarities in phenotype between KI and Fmr1 knockout mice and suggest that the decreased concentration of FMRP contributes to the phenotype in young adult KI mice.
Published by Elsevier Inc.
Figures
References
-
- Aziz M, et al. Clinical features of boys with fragile X premutations and intermediate alleles. American Journal of Medical Genetics Part B-Neuropsychiatric Genetics. 2003;121B:119–127. - PubMed
-
- Bontekoe CJM, et al. Instability of a (CGG)(98) repeat in the Fmr1 promoter. Human Molecular Genetics. 2001;10:1693–1699. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
