

The Ca2+ channel beta4c subunit interacts with heterochromatin protein 1 via a PXVXL binding motif
- PMID: 21220418
- PMCID: PMC3058963
- DOI: 10.1074/jbc.M110.187864
The Ca2+ channel beta4c subunit interacts with heterochromatin protein 1 via a PXVXL binding motif
Abstract
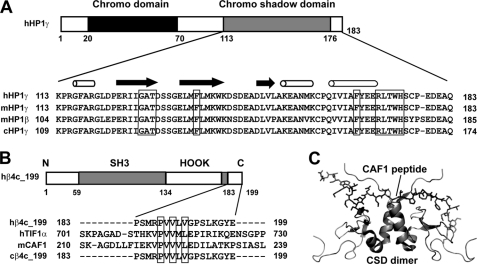
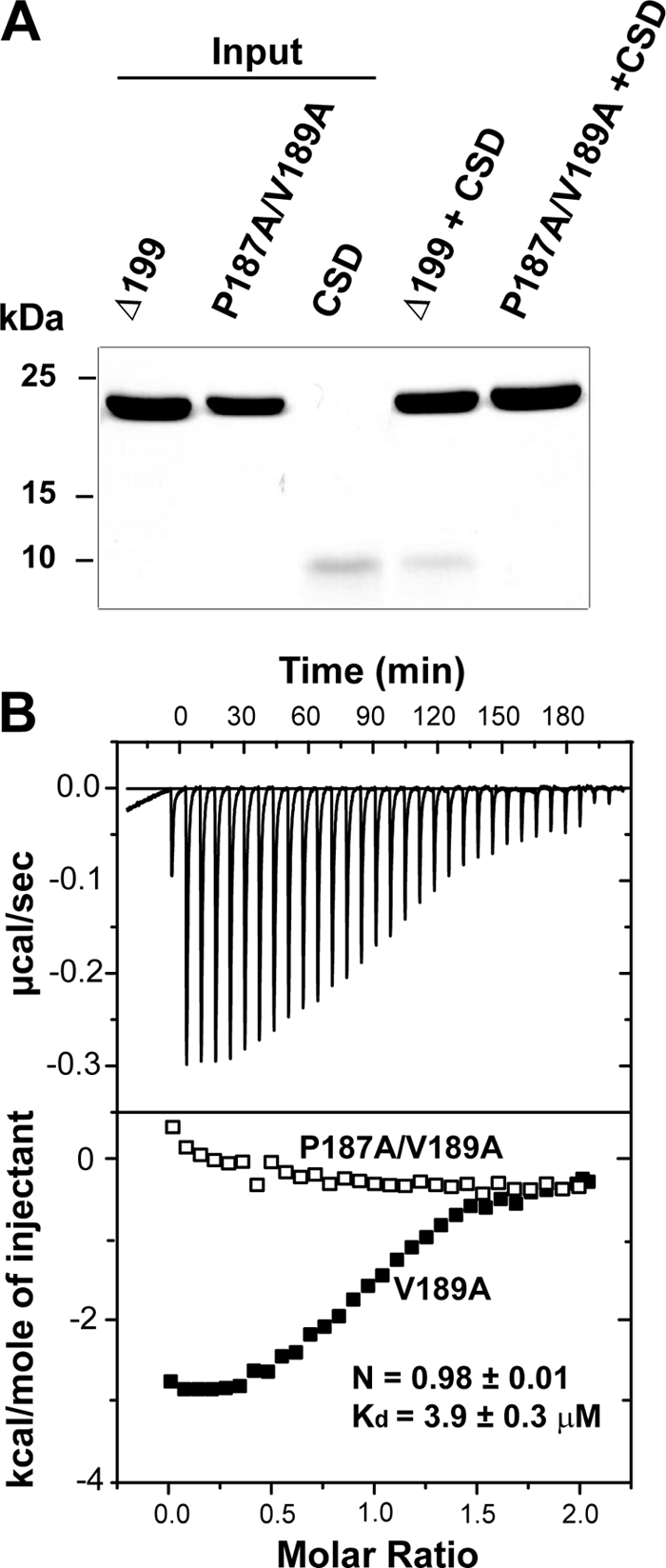
The β subunits of voltage-gated Ca(2+) channels are best known for their roles in regulating surface expression and gating of voltage-gated Ca(2+) channel α(1) subunits. Recent evidence, however, indicates that these proteins have a variety of Ca(2+) channel-independent functions. For example, on the molecular level, they regulate gene expression, and on the whole animal level, they regulate early cell movements in zebrafish development. In the present study, an alternatively spliced, truncated β4 subunit (β4c) is identified in the human brain and shown to be highly expressed in nuclei of vestibular neurons. Pull-down assays, nuclear magnetic resonance, and isothermal titration calorimetry demonstrate that the protein interacts with the chromo shadow domain (CSD) of heterochromatin protein 1γ. Site-directed mutagenesis reveals that the primary CSD interaction occurs through a β4c C-terminal PXVXL consensus motif, adding the β4c subunit to a growing PXVXL protein family with epigenetic responsibilities. These proteins have multiple nuclear functions, including transcription regulation (TIF1α) and nucleosome assembly (CAF1). An NMR-based two-site docking model of β4c in complex with dimerized CSD is presented. Possible roles for the interaction are discussed.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
