

Purinergic signaling in embryonic and stem cell development
- PMID: 21222015
- PMCID: PMC11114541
- DOI: 10.1007/s00018-010-0614-1
Purinergic signaling in embryonic and stem cell development
Abstract
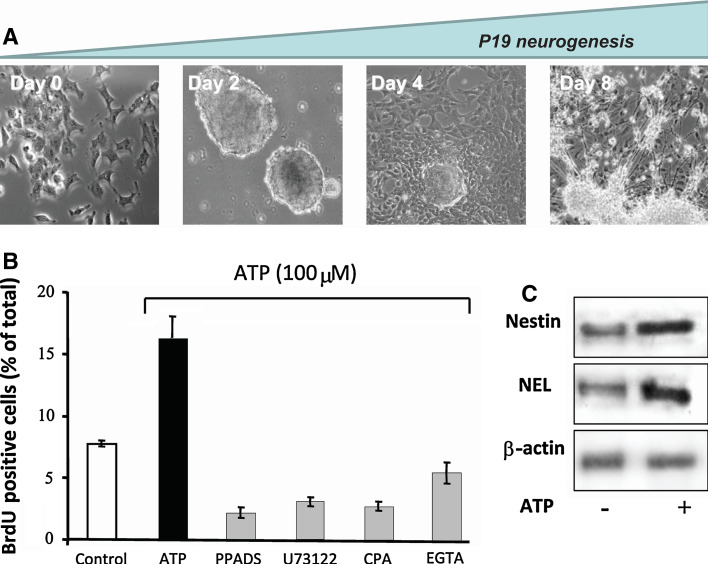
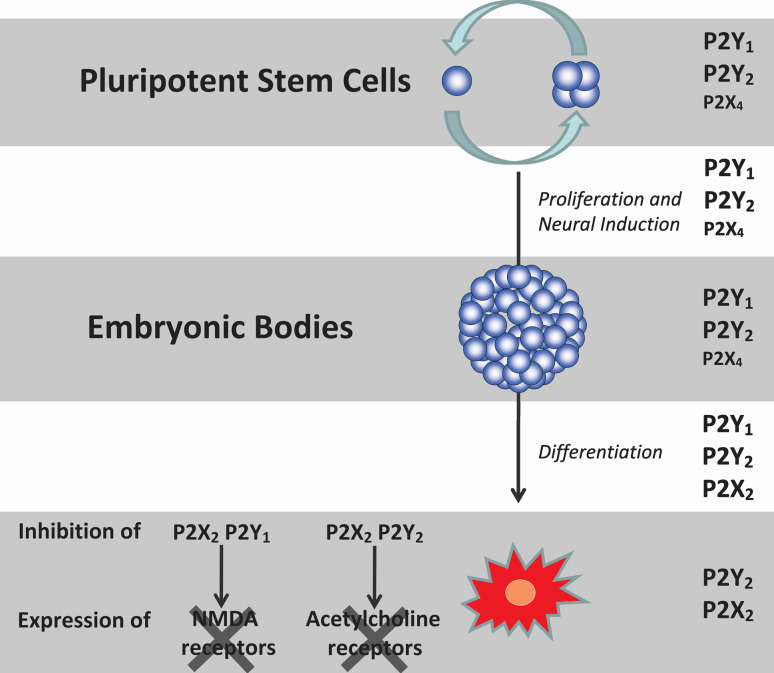
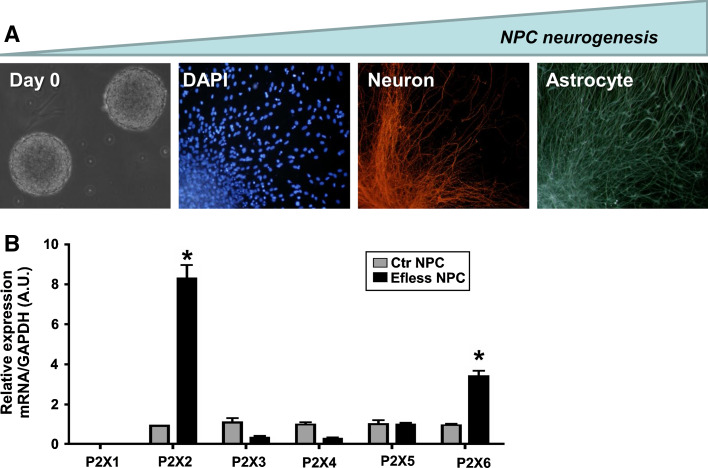
Nucleotides are of crucial importance as carriers of energy in all organisms. However, the concept that in addition to their intracellular roles, nucleotides act as extracellular ligands specifically on receptors of the plasma membrane took longer to be accepted. Purinergic signaling exerted by purines and pyrimidines, principally ATP and adenosine, occurs throughout embryologic development in a wide variety of organisms, including amphibians, birds, and mammals. Cellular signaling, mediated by ATP, is present in development at very early stages, e.g., gastrulation of Xenopus and germ layer definition of chick embryo cells. Purinergic receptor expression and functions have been studied in the development of many organs, including the heart, eye, skeletal muscle and the nervous system. In vitro studies with stem cells revealed that purinergic receptors are involved in the processes of proliferation, differentiation, and phenotype determination of differentiated cells. Thus, nucleotides are able to induce various intracellular signaling pathways via crosstalk with other bioactive molecules acting on growth factor and neurotransmitter receptors. Since normal development is disturbed by dysfunction of purinergic signaling in animal models, further studies are needed to elucidate the functions of purinoceptor subtypes in developmental processes.
Figures
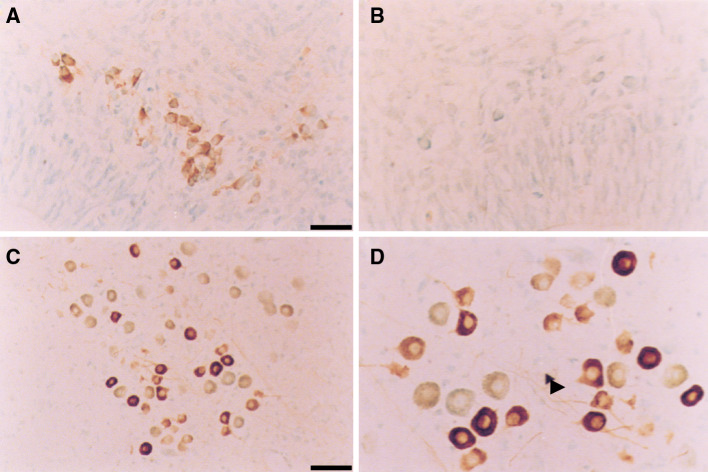
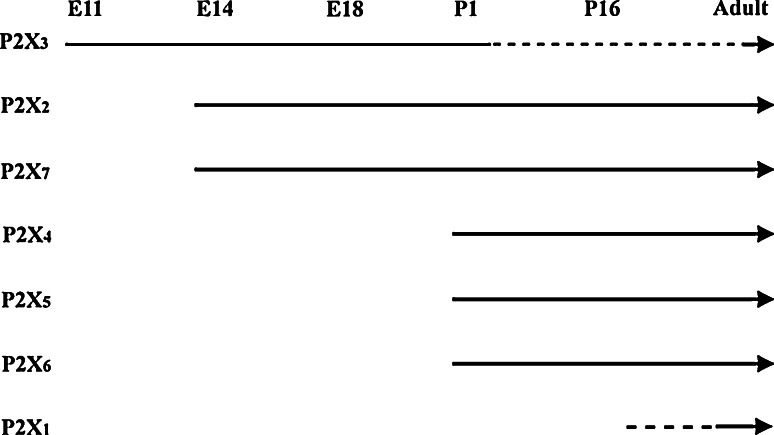
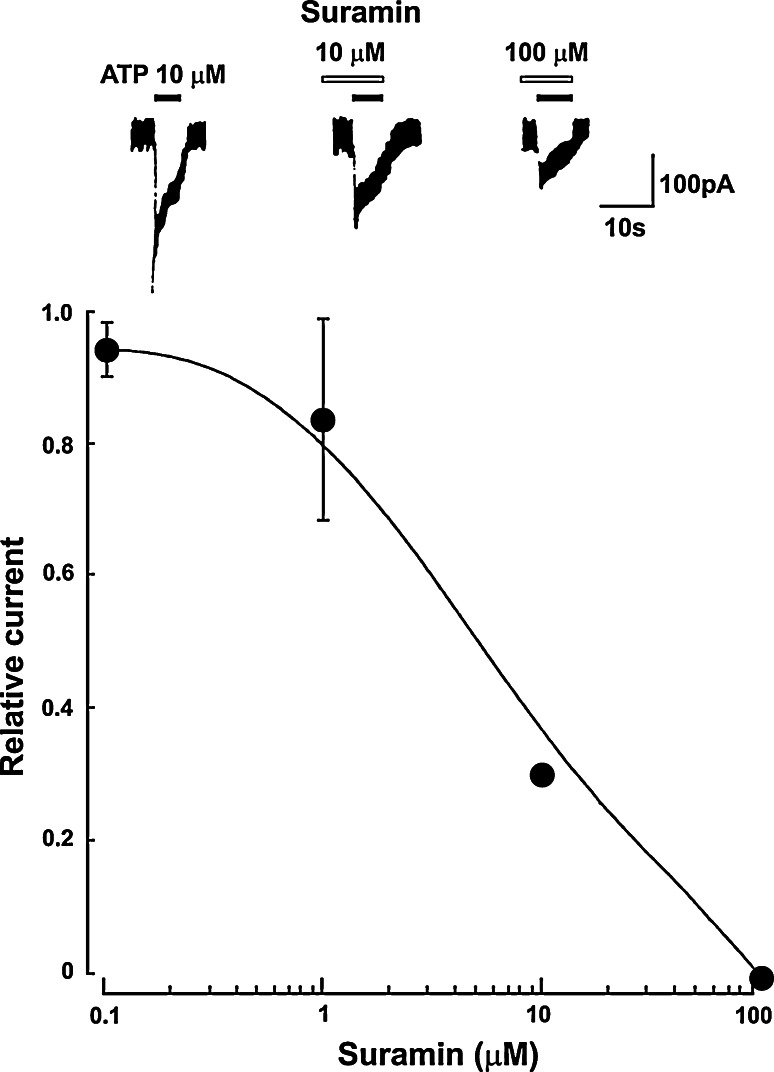
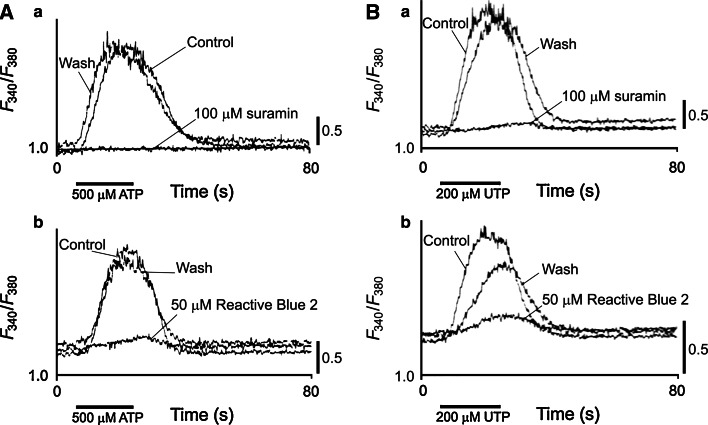
Similar articles
-
Purinergic signalling and development of the autonomic nervous system.Auton Neurosci. 2015 Sep;191:67-77. doi: 10.1016/j.autneu.2015.04.009. Epub 2015 Apr 30. Auton Neurosci. 2015. PMID: 25953245 Review.
-
Purines as potential morphogens during embryonic development.Purinergic Signal. 2012 Sep;8(3):503-21. doi: 10.1007/s11302-012-9290-y. Epub 2012 Jan 25. Purinergic Signal. 2012. PMID: 22270538 Free PMC article. Review.
-
Purinergic signaling in the retina: From development to disease.Brain Res Bull. 2019 Sep;151:92-108. doi: 10.1016/j.brainresbull.2018.10.016. Epub 2018 Nov 17. Brain Res Bull. 2019. PMID: 30458250 Free PMC article. Review.
-
Introduction to Purinergic Signaling.Methods Mol Biol. 2020;2041:1-15. doi: 10.1007/978-1-4939-9717-6_1. Methods Mol Biol. 2020. PMID: 31646477
-
Purinergic receptors in neurogenic processes.Brain Res Bull. 2019 Sep;151:3-11. doi: 10.1016/j.brainresbull.2018.12.013. Epub 2018 Dec 26. Brain Res Bull. 2019. PMID: 30593881 Review.
Cited by
-
Potentiation of temozolomide antitumor effect by purine receptor ligands able to restrain the in vitro growth of human glioblastoma stem cells.Purinergic Signal. 2015 Sep;11(3):331-46. doi: 10.1007/s11302-015-9454-7. Epub 2015 May 15. Purinergic Signal. 2015. PMID: 25976165 Free PMC article.
-
PKC-dependent ERK phosphorylation is essential for P2X7 receptor-mediated neuronal differentiation of neural progenitor cells.Cell Death Dis. 2013 Aug 1;4(8):e751. doi: 10.1038/cddis.2013.274. Cell Death Dis. 2013. PMID: 23907465 Free PMC article.
-
Impairment of adenosine signaling disrupts early embryo development: unveiling the underlying mechanisms.Front Pharmacol. 2024 Jan 19;14:1328398. doi: 10.3389/fphar.2023.1328398. eCollection 2023. Front Pharmacol. 2024. PMID: 38313072 Free PMC article.
-
Purinergic signaling: Diverse effects and therapeutic potential in cancer.Front Oncol. 2023 Jan 18;13:1058371. doi: 10.3389/fonc.2023.1058371. eCollection 2023. Front Oncol. 2023. PMID: 36741002 Free PMC article. Review.
-
Purinergic Signaling Modulates Survival/Proliferation of Human Dental Pulp Stem Cells.J Dent Res. 2019 Feb;98(2):242-249. doi: 10.1177/0022034518807920. Epub 2018 Nov 1. J Dent Res. 2019. PMID: 30383477 Free PMC article.
References
-
- Burnstock G. Physiology and pathophysiology of purinergic neurotransmission. Physiol Rev. 2007;87:659–797. - PubMed
-
- Kupitz Y, Atlas D. A putative ATP-activated Na + channel involved in sperm-induced fertilization. Science. 1993;261:484–486. - PubMed
-
- Smith R, Koenig C, Pereda J. Adenosinetriphosphatase (Mg-ATPase) activity in the plasma membrane of preimplantation mouse embryo as revealed by electron microscopy. Anat Embryol (Berl) 1983;168:455–466. - PubMed
-
- Ishikawa T, Seguchi H. Localization of Mg++-dependent adenosine triphosphatase and alkaline phosphatase activities in the postimplantation mouse embryos in day 5 and 6. Anat Embryol (Berl) 1985;173:7–11. - PubMed
-
- Foresta C, Rossato M, Di Virgilio F. Extracellular ATP is a trigger for the acrosome reaction in human spermatozoa. J Biol Chem. 1992;267:1–5. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
