

Hippocampal microcircuit dynamics probed using optical imaging approaches
- PMID: 21224219
- PMCID: PMC3090592
- DOI: 10.1113/jphysiol.2010.202184
Hippocampal microcircuit dynamics probed using optical imaging approaches
Abstract
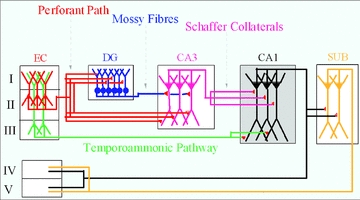
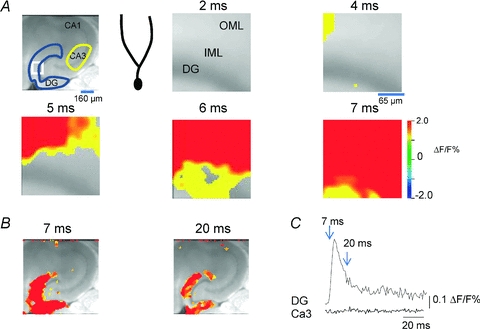
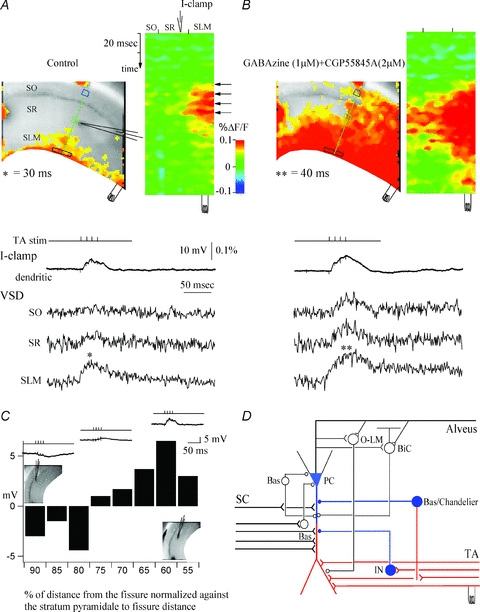
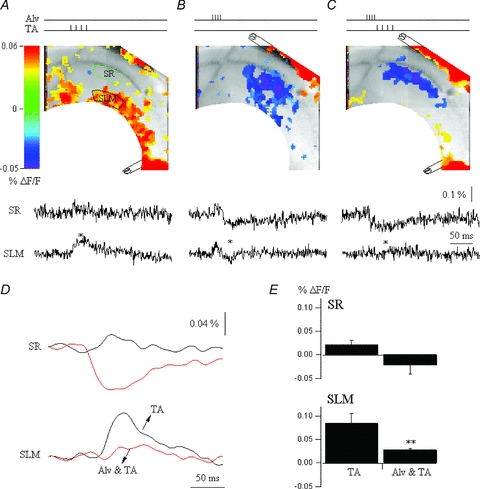
Mammalian cortical structures are endowed with the capacity for plasticity, which emerges from a combination of the dynamics of circuit connectivity and function, and the intrinsic function of the neurons within the circuit. However, this capacity is accompanied by a significant risk: the capability to generate seizure discharges is also a property of all mammalian cortices. How do cortical circuits reconcile the requirement to maintain plasticity, but at the same time control seizure initiation? These issues come into particular focus in the hippocampus. The hippocampus is one of the main plasticity engines in the brain, and is also a structure frequently implicated in the generation of epileptic seizures, with temporal lobe epilepsy constituting the most prevalent form of epilepsy in the adult population. One aspect of hippocampal circuitry that is particularly prominent is its intimate interconnections with the entorhinal cortex. These interconnections create a number of excitatory synaptic loops within the limbic system, which, in addition to being important in cognitive function, can support reentrant activation and seizure generation. In the present review, using optical imaging approaches to elucidate circuit processing at high temporal and spatial resolution, we examine how two targets of entorhinal cortical input within the hippocampus, the dentate gyrus and area CA1, regulate these synaptic pathways in ways that can maintain functions important in generation of normal activity patterns, but that dampen the ability of these inputs to generate seizure discharges.
Figures
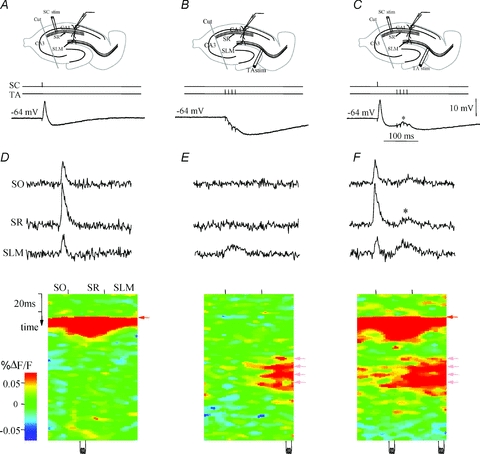
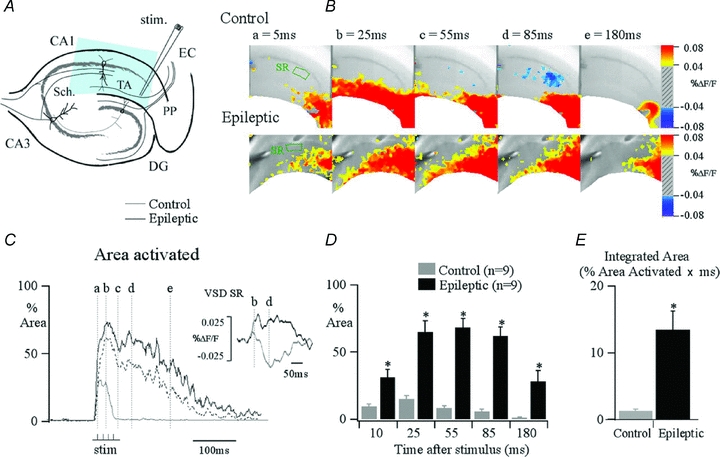
References
-
- Brooks-Kayal AR, Shumate MD, Hong Y, Rikhter TY, Coulter DA. Selective changes in single cell GABAA receptor subunit expression correlate with altered function in epileptic hippocampus. Nat Med. 1998;4:1166–1172. - PubMed
-
- Brun VH, Otnaess MK, Molden S, Steffenach HA, Witter MP, Moser MB, Moser EI. Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry. Science. 2002;296:2243–2246. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous