

A three-dimensional multiscale model for gas exchange in fruit
- PMID: 21224337
- PMCID: PMC3046576
- DOI: 10.1104/pp.110.169391
A three-dimensional multiscale model for gas exchange in fruit
Abstract
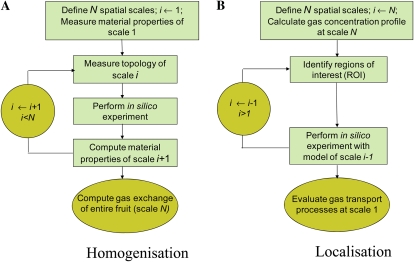
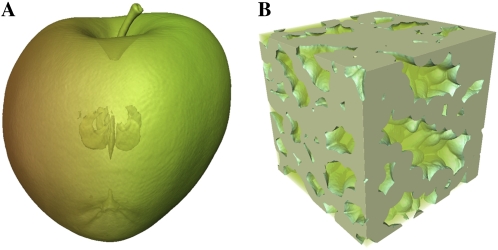
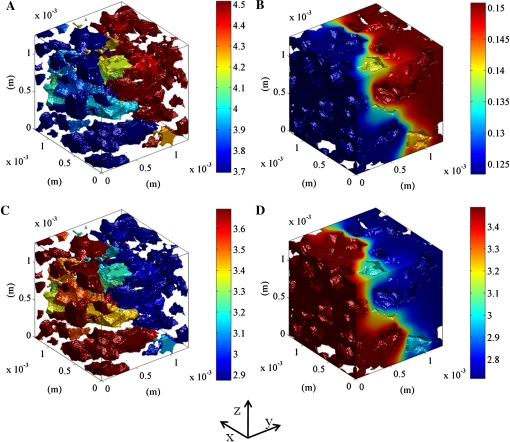
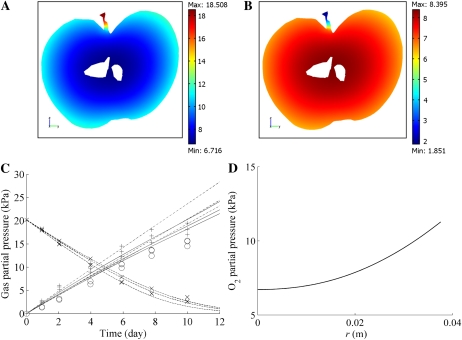
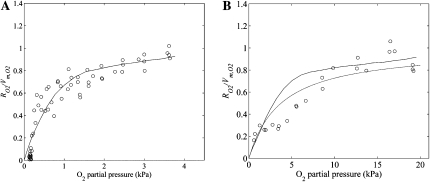
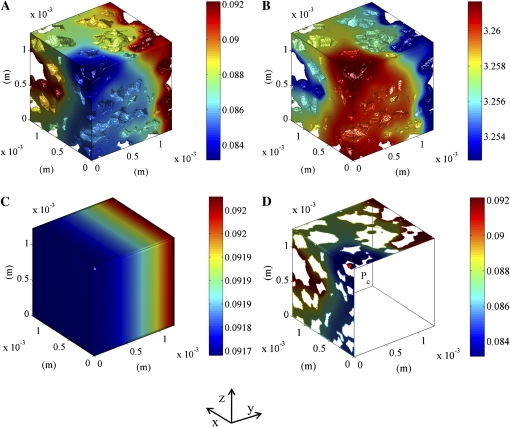
Respiration of bulky plant organs such as roots, tubers, stems, seeds, and fruit depends very much on oxygen (O2) availability and often follows a Michaelis-Menten-like response. A multiscale model is presented to calculate gas exchange in plants using the microscale geometry of the tissue, or vice versa, local concentrations in the cells from macroscopic gas concentration profiles. This approach provides a computationally feasible and accurate analysis of cell metabolism in any plant organ during hypoxia and anoxia. The predicted O2 and carbon dioxide (CO2) partial pressure profiles compared very well with experimental data, thereby validating the multiscale model. The important microscale geometrical features are the shape, size, and three-dimensional connectivity of cells and air spaces. It was demonstrated that the gas-exchange properties of the cell wall and cell membrane have little effect on the cellular gas exchange of apple (Malus×domestica) parenchyma tissue. The analysis clearly confirmed that cells are an additional route for CO2 transport, while for O2 the intercellular spaces are the main diffusion route. The simulation results also showed that the local gas concentration gradients were steeper in the cells than in the surrounding air spaces. Therefore, to analyze the cellular metabolism under hypoxic and anoxic conditions, the microscale model is required to calculate the correct intracellular concentrations. Understanding the O2 response of plants and plant organs thus not only requires knowledge of external conditions, dimensions, gas-exchange properties of the tissues, and cellular respiration kinetics but also of microstructure.
Figures
References
-
- Aalto T, Juurola E. (2002) A three-dimensional model of CO2 transport in airspaces and mesophyll cells of a silver birch leaf. Plant Cell Environ 25: 1399–1409
-
- Bidel LPR, Renault P, Pagès L, Rivière LM. (2000) Mapping meristem respiration of Prunus persica (L.) Batsch seedlings: potential respiration of the meristems, O2 diffusional constraints and combined effects on root growth. J Exp Bot 51: 755–768 - PubMed
-
- Borisjuk L, Rolletschek H. (2009) The oxygen status of the developing seed. New Phytol 182: 17–30 - PubMed
-
- Colmer TD, Voesenek LACJ. (2009) Flooding tolerance: suites of plant traits in variable environments. Funct Plant Biol 36: 665–681 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
