

The Cullin-RING E3 ubiquitin ligase CRL4-DCAF1 complex dimerizes via a short helical region in DCAF1
- PMID: 21226479
- PMCID: PMC3072279
- DOI: 10.1021/bi101749s
The Cullin-RING E3 ubiquitin ligase CRL4-DCAF1 complex dimerizes via a short helical region in DCAF1
Abstract
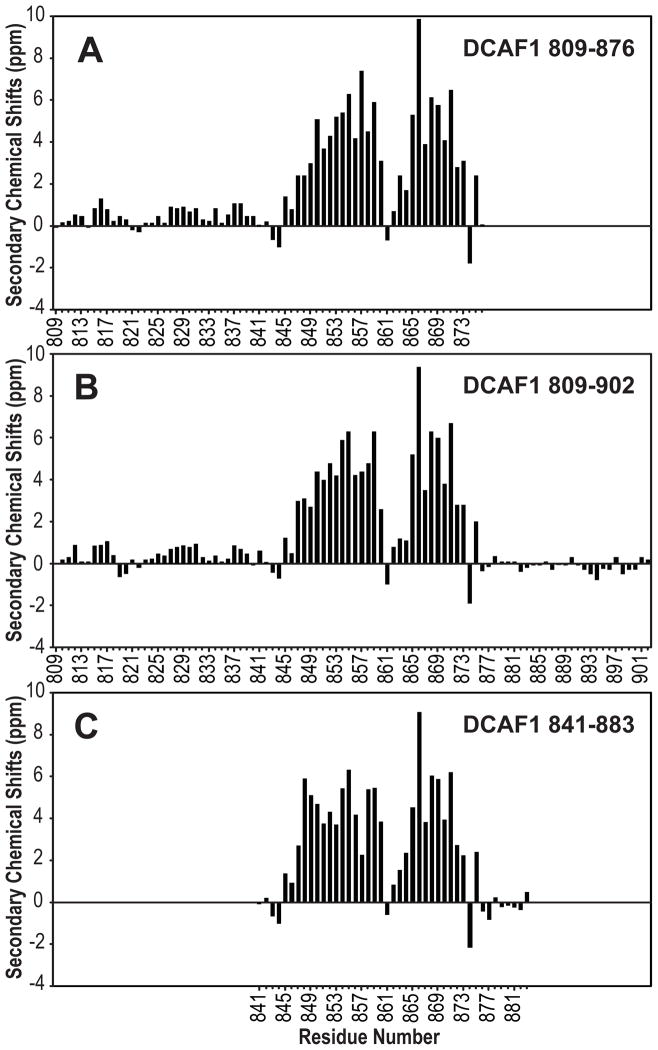
The cullin4A-RING E3 ubiquitin ligase (CRL4) is a multisubunit protein complex, comprising cullin4A (CUL4), RING H2 finger protein (RBX1), and DNA damage-binding protein 1 (DDB1). Proteins that recruit specific targets to CRL4 for ubiquitination (ubiquitylation) bind the DDB1 adaptor protein via WD40 domains. Such CRL4 substrate recognition modules are DDB1- and CUL4-associated factors (DCAFs). Here we show that, for DCAF1, oligomerization of the protein and the CRL4 complex occurs via a short helical region (residues 845-873) N-terminal to DACF1's own WD40 domain. This sequence was previously designated as a LIS1 homology (LisH) motif. The oligomerization helix contains a stretch of four Leu residues, which appear to be essential for α-helical structure and oligomerization. In vitro reconstituted CRL4-DCAF1 complexes (CRL4(DCAF1)) form symmetric dimers as visualized by electron microscopy (EM), and dimeric CRL4(DCAF1) is a better E3 ligase for in vitro ubiquitination of the UNG2 substrate compared to a monomeric complex.
Figures






References
-
- Ciechanover A, Finley D, Varshavsky A. Ubiquitin dependence of selective protein degradation demonstrated in the mammalian cell cycle mutant ts85. Cell. 1984;37:57–66. - PubMed
-
- Ciechanover A, Finley D, Varshavsky A. The ubiquitin-mediated proteolytic pathway and mechanisms of energy-dependent intracellular protein degradation. J Cell Biochem. 1984;24:27–53. - PubMed
-
- Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–479. - PubMed
-
- Pickart CM. Mechanisms underlying ubiquitination. Annu Rev Biochem. 2001;70:503–533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
