

doi: 10.1103/PhysRevLett.105.218101.
Epub 2010 Nov 17.
Factors governing fibrillogenesis of polypeptide chains revealed by lattice models
Affiliations
- PMID: 21231356
- PMCID: PMC11298782
- DOI: 10.1103/PhysRevLett.105.218101
Item in Clipboard
Factors governing fibrillogenesis of polypeptide chains revealed by lattice models
Phys Rev Lett.
.
Abstract
Using lattice models we explore the factors that determine the tendencies of polypeptide chains to aggregate by exhaustively sampling the sequence and conformational space. The morphologies of the fibril-like structures and the time scales (τ(fib)) for their formation depend on a balance between hydrophobic and Coulomb interactions. The extent of population of an ensemble of N* structures, which are fibril-prone structures in the spectrum of conformations of an isolated protein, is the major determinant of τ(fib). This observation is used to determine the aggregating sequences by exhaustively exploring the sequence space, thus providing a basis for genome wide search of fragments that are aggregation prone.
Figures
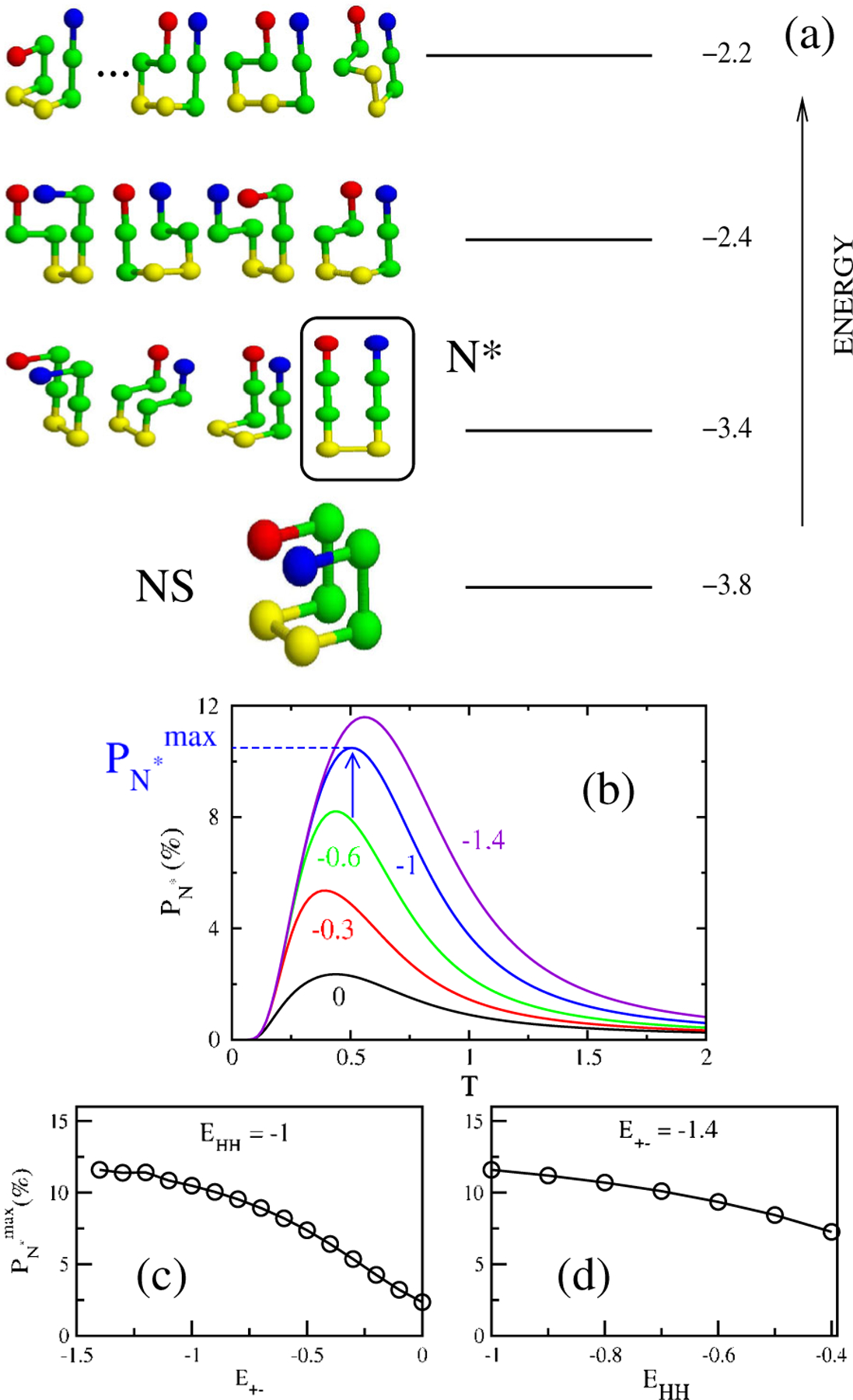
(a) Spectrum of energies and low energy structures of the monomer
sequence , , , + and − are in green, yellow, blue, and
red, respectively. We set and . There are 1831 possible conformations that are
spread among 17 possible energy values. The conformations in the first excited
state represent the ensemble of structures and the conformation that coincides with the peptide
state in the fibril [see Fig. 2(a)] is
enclosed in a box. (b) The probability of populating the structure in the box in (a)
as a function of for and −1.4 keeping
. The arrow indicates , where . Dependence of on for (c), and on for (d).
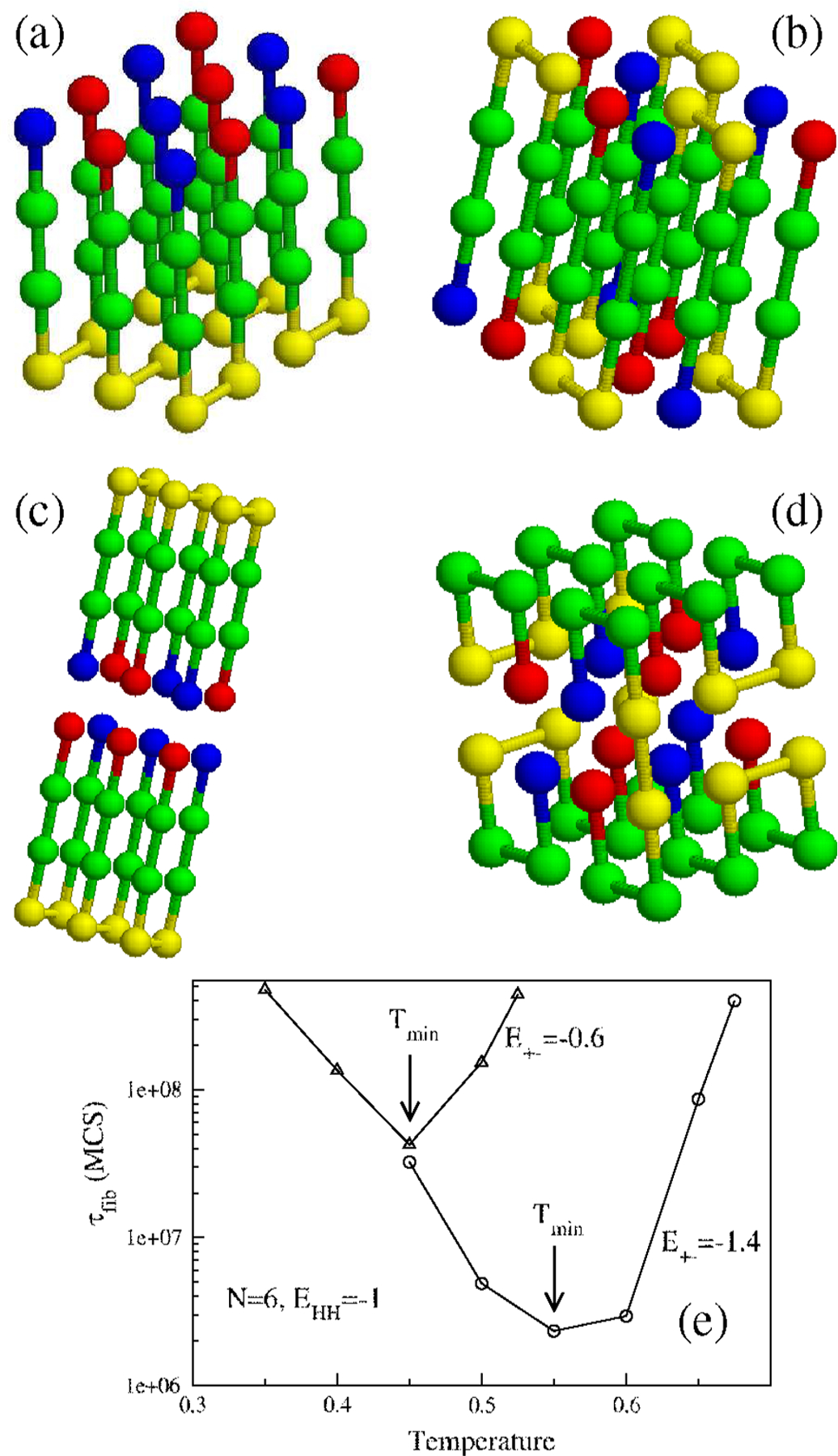
(a) The lowest-energy fibril structure for and . (b) Same as in (a) but with
. (c) Double-layer structure for
but with . (d) For and the fibril structure is entirely altered. (e)
Temperature dependence of for (circles) and (triangles). and . Arrows show the temperatures at which the
fibril formation is fastest.
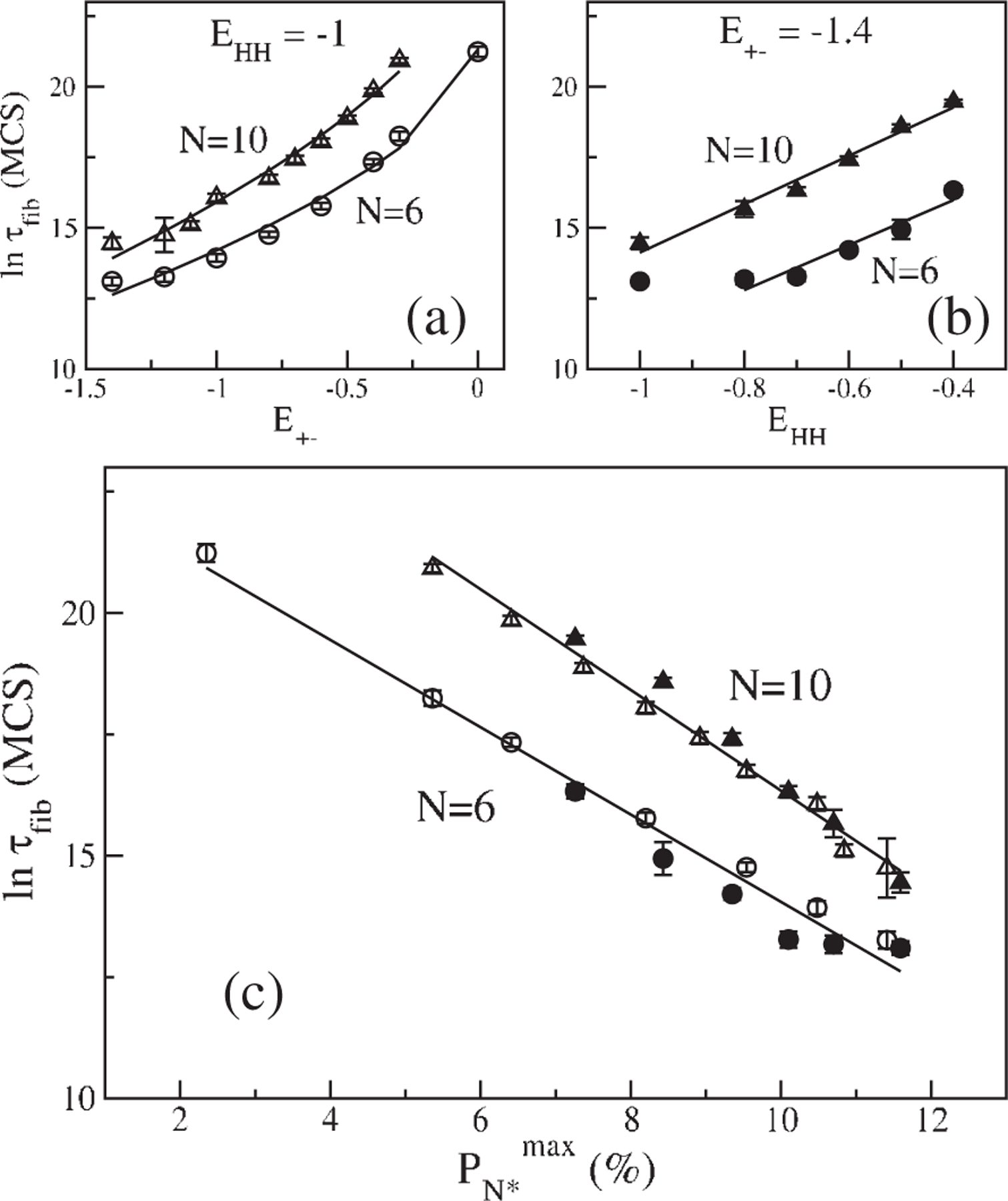
(a) Dependence of on for (circles) and (triangles) with . The solid curves are fits to
, where . and and and for and 10 , respectively. (b) Dependence of
on with hold constant for (solid circle) and (solid triangles). Lines are fits
and for and 10 , respectively. For
the first point is excluded from fitting. (c) Dependence of
on for and 10 . Symbols are the same as in (a) and (b)
is measured in MCS and
in percent. The correlation coefficient for all
fits .
References
-
- (a) Otzen DE, Kristensen O, and Oliveberg M, Proc. Natl. Acad. Sci. U.S.A. 97, 9907 (2000); - PMC - PubMed
- (b) Massi F, Klimov D, Thirumalai D, and Straub JE, Prot. Sci. 11, 1639 (2002); - PMC - PubMed
- (c) Chiti F, Calamai M, Taddei N, Stefani M, Ramponi G, and Dobson CM, Proc. Natl. Acad. Sci. U.S.A. 99, 16419 (2002); - PMC - PubMed
- (d) West MW, Wang WX, Patterson J, Mancias JD, Beasley JR, and Hecht MH, Proc. Natl. Acad. Sci. U.S.A. 96, 11211 (1999); - PMC - PubMed
- (e) Kallberg Y, Gustafsson M, Persson B, Thyberg J, and Johansson J, J. Biol. Chem. 276, 12945 (2001); - PubMed
- (f) Dima RI and Thirumalai D, Biophys. J. 83, 1268 (2002); - PMC - PubMed
- (g) Gazit E, FASEB monographs 16, 77 (2002). - PubMed
-
- (a) Klimov DK and Thirumalai D, Structure 11, 295 (2003); - PubMed
- (b) Bellesia G and Shea JE, Biophys. J. 96, 875 (2009); - PMC - PubMed
- (c) de la Paz ML, de Mori GMS, Serrano L, and Colombo G, J. Mol. Biol 349, 583 (2005); - PubMed
- (d) Li DW, Mohanty S, Irback A, and Huo SH, PLoS Comput. Biol 4, e1000238 (2008). - PMC - PubMed
-
- (a) Klimov DK and Thirumalai D, J. Chem. Phys. 109, 4119 (1998);
- (b) Shakhnovich E, Chem. Rev 106, 1559 (2006); - PMC - PubMed
- (c) Gupta P, Hall CK, and Voegler AC, Protein Sci 7, 2642 (1998); - PMC - PubMed
- (d) Dima RI and Thirumalai D, Protein Sci 11, 1036 (2002); - PMC - PubMed
- (e) Maiti M, Rao M, and Sastry S, Eur. Phys. J. E 32, 217 (2010); - PubMed
- Cieplak M, Hoang TX, and Li MS, Phys. Rev. Lett 83, 1684 (1999);
- Li MS, Klimov DK, and Thirumalai D, Phys. Rev. Lett 93, 268107 (2004); - PubMed
- Li MS and Cieplak M, Phys. Rev. E 59, 970 (1999);
- Kouza M, Li MS, O’Brien EP, Hu C-K, and Thirumalai D, J. Phys. Chem. A 110, 671 (2006). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources